Created by Kamila Klavíková
Částečné zápisky z přednášky Vývojové biologie na PřF UK.
Spermie ježovky mají na sobě bindinové ligandy, které se odhalí po vylití akrozomálního váčku, a vajíčko má bindinové receptory. Vážou se k sobě jako zámek a klíč. U mořských živočichů je důležitá tato tvrdá selekce, aby nebyla zmařená celá dávka spermií na špatný druh.
Akrozomální váček obsahuje 26S proteazomální komplex, který slouží k degradaci ubiquitinovaných proteinů. Váček se vylije během akrozomální reakce. Ubiquitin se nachází i na vnějšku zony pellucidy. Ubiquintylace je druhově specifická. Tyto ubiquintylované proteiny degraduje 26S proteazomální komplex. Zablokování proteazomálního komplexu brání oplození u zona pellucida intaktních savců.
Ubiquitin se skládá z 76 aminokyselin. V rámci postranslančních modifikcí určitých proteinů (určených k degradaci) se pomocí UBA a UBC enzymů kovalentně váže na lyzinové zbytky. Ubiquitin sám obsahuje 7 lyzinových zbytků – možnost tvorby polymerů (di, tri, tetra nebo poly). 26S proteazomální komplex rozpoznává ubiquitinylované proteiny a degraduje je.
#otazka Jak se tvoří buňky blastému? Kde se berou?
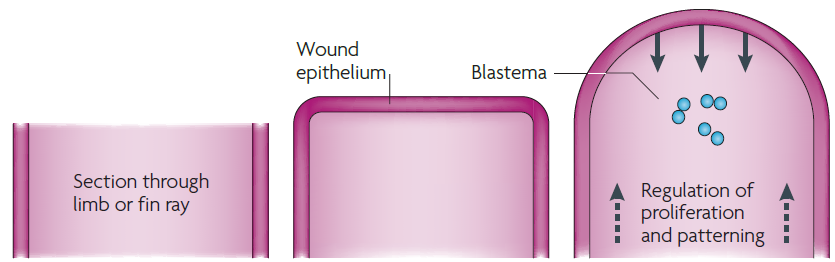
Blastém je masa buněk schopná růstu a regenerace v orgány nebo části těla.
Po té, co došlo k amputaci končetiny, se rána zakryje vrstvou pokožky. Tato pokožka se přemění v apikální ektodermální čepičku (AEC). Buňky dediferencují, proliferencují, diferencují.
Vývoj končetiny mloka během regenerace je podobný vývoji končetiny během embryonálního vývoje. Vytváří se apikální ektodermální čepička (apical ectodermal cap), což je obdoba apikálního ektodermálního výběžku (AER). K tvorbě blastému přispívá řada tkání. Uchovávájí si svoji původní identitu i přes dediferenciaci a následnou proliferaci.
Při akrozomální reakci dojde k vylití akrozomálního váčku s lytickými enzymy, které začnou rozpouštět extracelulární obal (vitelinní membránu nebo zonu pellucidu). U ježovky je tato reakce závislá na koncentraci Ca2+.
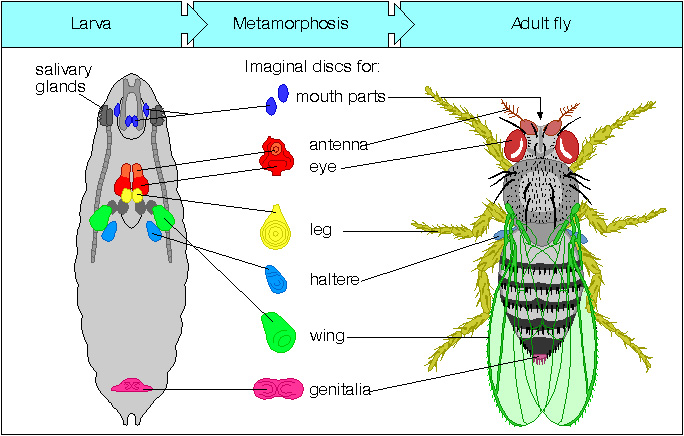
Larva obsahuje dvě populace buněk – buňky vlastní larvální tkáně a larválních orgánů a buňky budoucích tkání a orgánů dospělce – imaginální buňky.
Imaginální buňky jsou nediferenciované. Během lárválního vývoje proliferují. K diferenciaci dochází až během metamorfózy.
Larvální tkáň přispívá jen minimálně k tvorbě těla dospělce. Zachovává se jen část nervové soustavy. Larvální tkáň během metamorfózy podstupuje programovanou buněčnou smrt.
Tělo dospělce je vytvořeno z imaginálních buněk.
Regeneracce je obnovení vývoje během dospělosti. Cílem regenerace je obnova poškozených nebo chybějících tkání.
Epimorfóza je charakterizována tvorbou regeneračního blastému. Buňky stávající tkáně dediferenciují, proliferují a následně se diferenciují.
Př. končetiny a ocas mloka, ocas žabích pulců, ploutve ryb, ostnokožci, měkýšši, ploštěnci.
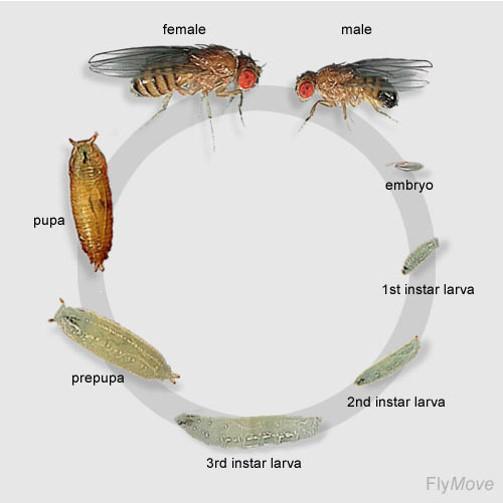
na příkladu Drosophila melanogaster
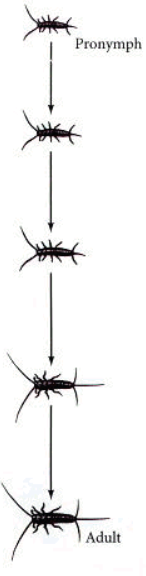
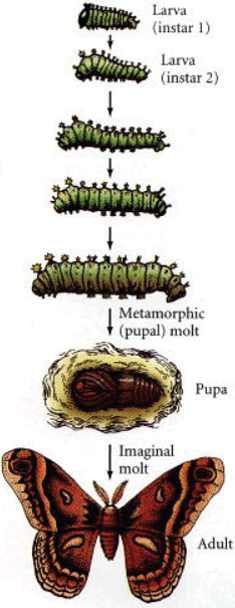
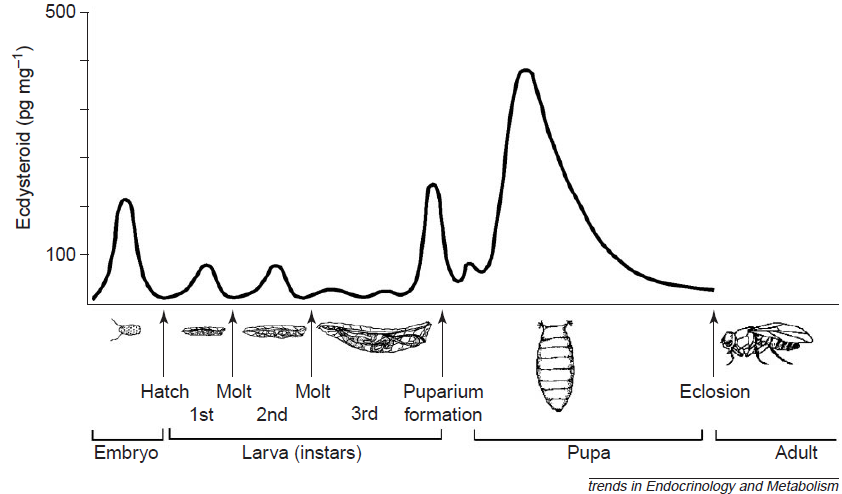
Vývoj přes tři larvální stádia – instary. Larva třetího instaru se zakuklí. V kukle proběhne metamorfóza. Z kukly se vylíhne dospělý jedinec – imago. Celý vývoj trvá zhruba dva týdny.
Po rychlém bloku nastává kortikální reakce. Za zásob se vylévají Ca2+. Pod membránou vajíčka dojde k fúzi váčku obsahujících enzymy s vitelinní membránou (u ježovky). V nich se nachází serinové proteázy, které odštěpí serinové proteázy, dále mukopolysacharidy, které vážou vodu a vytvoří „vodní příkop“, též peroxidáza, která způsobí ztvrdnutí fertilizačního obalu, tím že propojí tyrosinové zbytky na vitelinní membráně, a hyelin, který vytvoří trvalý ochranný obal.
Při rychlém bloku dochází ke změnám membránového potenciálu. Po vniku první spermie do vajíčka se začne zvedat koncentrace Na+. Dojde k přepólování membrány a odmrštění spermií. Dlouhodobě to může být pro vajíčko nebezpečné. Po odmrštění se membránový potenciál navrátí do původního stavu.
Morfolaxe je přestavba tkání. Nedochází k proliferaci.
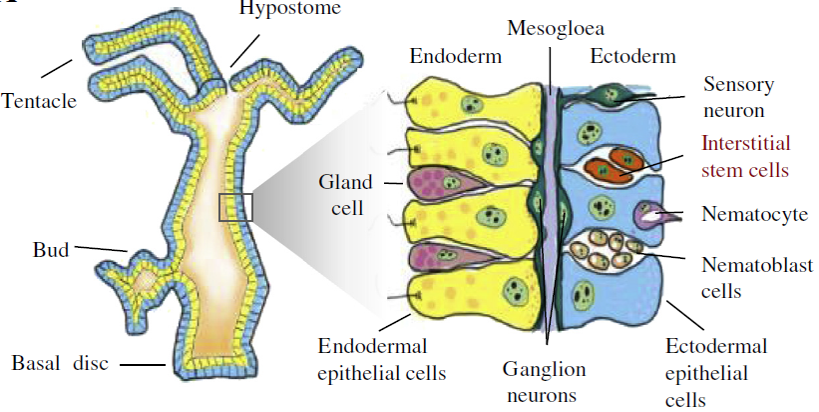
Tělo nezmara je tvořeno ektodermem a mezodermem. Ústní otvor s věncem chapadel (hypostom) je umístěn apikálně. Nožní terč je umístěn bazálně. Množí se pohlavně nebo pučením. Tělní buňky se neustále dělí a posouvají se podél apiko-bazální osy. Dělí se ekto- a endodermální epiteliální buňky a intersticiální kmenové buňky.
Za specifických podmínek může docházet i k epimorfóze.
Dělají se transplantační experimenty jako důkaz existence „organizátorů“. Po transplantaci z oblasti hypostomu vznikne sekundární osa s hypostomem – „hlavový organizátor“.
Pokud transplantujeme tkáň z oblasti bazálního disku, vznikne sekundární osa s bazálním diskem – „organizátor bazálního disku“. Dojde-li k transplantaci tkáně z oblasti hypostomu do blízkosti jiného hypostomu, sekundární osa nevznikne. Inhibiční signál zamezuje tvorbě dalších organizačních center.
Ihibice inzulínové dráhy vede k prodloužení života.
inzulín (signál přichází v závislosti na vnějších podmínkách) → receptor pro inzulín → přenos signálu do jádra → aktivace transkripce
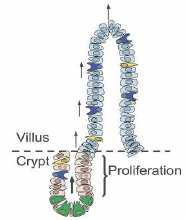 | 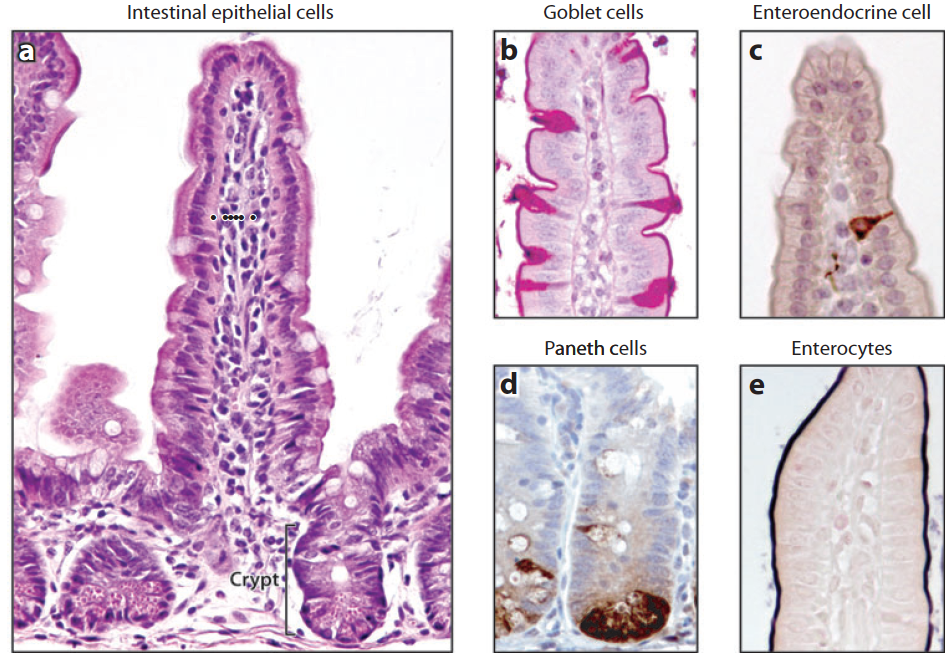 |
Výstelka střeva je tvořená jednovrstevným epitelem zřaseným do útvarů nazývaných klky a krypty.
Střevní epitel tvoří mnoho typů buněk. Nalezneme tu:
Mimo to jsou přítomné další typy buněk, jež tvoří součást imunitního systému sliznice a moc se o nich neví (tuft cells, M cells, cup cells).
Dále jsou přítomné kmenové buňky. Bez nich by nebyla možná úplná obnova střevního epitelu každých 4 až 5 dní. Leží na dně každé krypty, kde jich je jen několik. Jsou obklopené Panethovými buňky, které pro ně vytváří nejvhodnější prostředí.
z řeckého meta – změn a morphe – forma
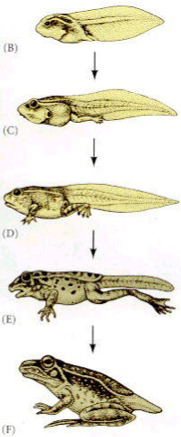
Metamorfóza je přeměna organismu z larvy na dospělce doprovázená morfologickou přestavbou těla. Další změny mohou probíhat na úrovni fyziologie, biochemi, etologie, ekologie.
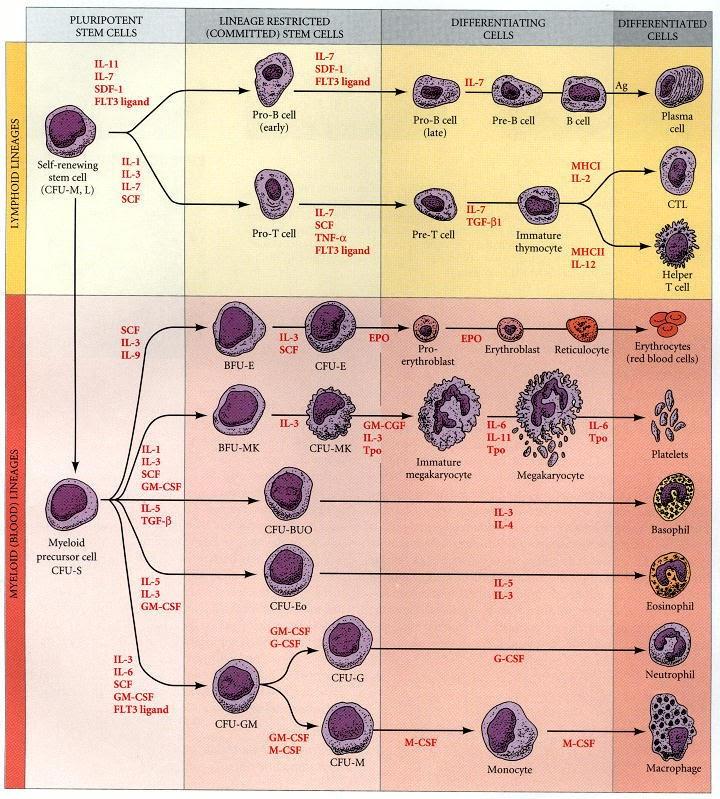
Později se vytváří hematopoetické buňky (HSC) v oblasti kolem aorty. Tyto buňky kolonizují emryonální játra.
V období kolem narození hematopoetické buňky kolonizují kostní dřeň – hlavní krvetvorný orgán dospělého organismu.
HSC dávají vzniknout všem typům buněk lymfoidní i myeloidní tkáně. Na začátku máme kmenovou buňku, která se jednak obnovuje, jednak dává vznik dalším typům kmenových buněk, jejichž osud je postupně specifikován, z multipotentních kmenových buněk se postupně stávájí unipotentní kmenové buňky.
Nejprve je spermie chemotaxí přitahovaná k vajíčku. Vajíčko vylučuje rozpustné molekuly.
Následně dojde k exocytóze akrozomálního váčku a vylití lytických enzymů (u savců nejprve vazba spermie k vajíčku). Spermie se naváže na extracelulární obal vajíčka (vitelinní membrána nebo zona pellucida).
Spermie (i s bičíkem, nikde ho neodhazuje) pronikne do vajíčka s pomocí enzymů. Plamatické membrány vajíčka a spermie splynou.
#otazka Proč by měly být biologicky nesmrtelné? Co to znamená být biologicky nesmrtelný?
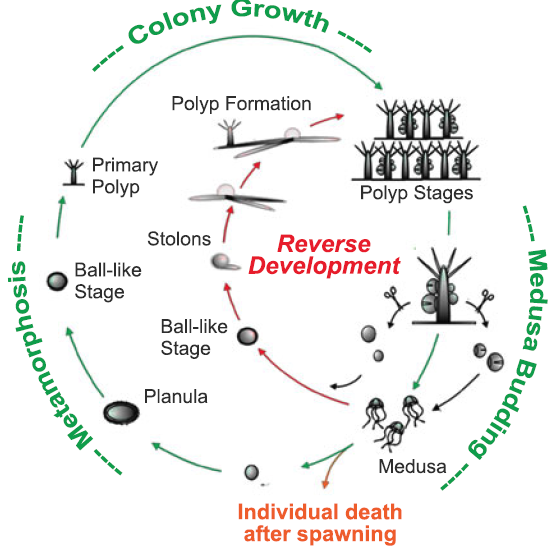
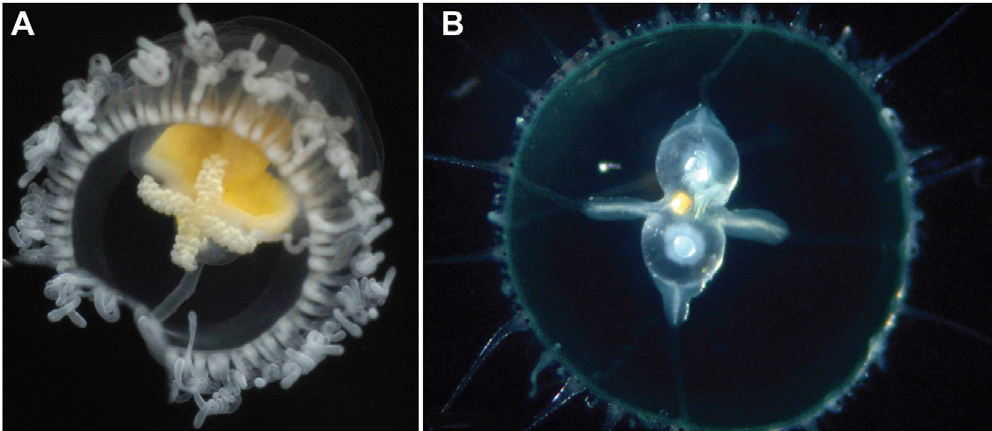
Některé druhy medůz jsou schopné přeměnit se z pohlavně dospělé medúzy zpět do stádia polypa. Děje se tak procesem transdiferenciace. Jsou teoreticky biologicky nesmrtelné.
V organismu se postupně hromadí nejrůznější poškození, jak na úrovni tkání, tak na úrovni genomu, zároveň klesá účinnost reparačních mechanismů. Organismus stárne.
Rozeznáváme dva základní typy obalů:
ovum
Vajíčko je samičí pohlavní buňka. V cytoplasmě obsahuje zásobní a ochranné látky, dále pak proteiny potřebné pro počáteční fáze zárodečného vývoje a obrovské množstí ribozomů, které umožňují translaci uložených RNA molekul. Z pohledu úspěšného oplození jsou podstatnou součástí vajíčka obaly.
Mimo cytoplasmatické membrány, kterou mají všechny buňky, obsahuje jako druhou vrstvu (obal) buď vitelinní membránu (ježovky), nebo zonu pellucidu (savci). Tento obal má regulační funkci, umožní navázání spermie na vajíčka a v ideálních případě průnik pouze jedné spermie. Dalším obalem je buď gelový obal (ježovky), nebo kumulární buňky (savci). Jejich funkce je opět regulační a ochranná.
Polyspermie je průnik dvou nebo více spermií do vajíčka. Spermie s sebou nosí centriolu, takže to vede ke vzniku více pólů dělícího vřeténka (4, 6). V důsledku toho vzniká fragmentované embryo s nedostatečnou genetickou informací. K polyspermii může například dojít při předčasné ovulaci, kdy má vajíčko slabou zonu pellucidu.
Hayflickův limit je kritická délka telomer, při které již buňka není schopná další reprodukce.
ESC
Embryonální kmenové buňky mají největší diferenciační potenciál. S manipulací s těmito buňkami jsou však spojené etické problémy. Abychom mohli ESC získat, je nutné zabít embryo. Existují postupy kultivace in vitro. Mohou být možné problémy s odvržením buněk příjemcem.
#otazka Jak se liší kompenzační regenerace od epimorfózy? Kde bere buňky?
Kompenzační regenerace je podobná epimorfóze, dochází k dediferenciaci, proliferaci a následné diferenciaci. Nemusí docházet k plné dediferenciaci. Netvoří se blastém.
Imaginální buňky jsou buňky budoucích tkání a orgánů dospělce.
Imaginální buňky dávají vzniknout kutikulárních útvarům dospělce. Př. křídla, končetiny, tykadla, ústní ústrojí, oči, genitálie.
Histoblasty dávají vzniknout tkáni zadečku.
Př. trávicí soustava, tracheální systém, slinné žlázy…
#otazka Jak probíhá splynutí spermie a „vajíčka“ v případě, že k průniku spermie dojde před dokončením oogeneze? Jak se oplození liší v závislosti na tom, v které fázi se oogeneze nachází?
Oplození představuje vznik nového organismu splynutím dvou gamet. Má několik klíčových fází.
Kapacitace je proces, při kterém se dokončí vývoj spermie. Ke kapacitaci dochází v pohlavním traktu samice.
Dojde k vylití K+ iontů, vtoku HCO3- iontů a podílí se na něm albumin.
Změní se vlastnosti spermie. Za prvé dojde k odstranění cholesterolu z membrány, takže v důsledku bude tekutá. Za druhé se aktivují protein-kinázy pro pohyb.
Stárnutí (senescence) je postupné zhoršování fyziologických funkcí nezbytných pro plodnost a přežití.
Hlavními aspekty jsou:
Telomery jsou speciální krátké repetetivní sekvence DNA, která se nachází na koncích chromozomu. Umožňují replikaci konců DNA beze ztráty genetického materiálu. Zároveň chrání DNA před degradací (aby se dala poznat od vadné). Telomeráza je reverzní transkriptáza. RNA využívá jako templát, podle kterého prodlužuje konce hromozomů. U dospělých jedinců není aktivní. Je aktivní v kmenových buňkách a v nádorových buňkách.
Při každém dělení dochází ke zkracování telomer, po dosažení určité kritické délky buňka není schopna další reprodukce (Hayflickův limit), stává se senescentní a posléze umírá. Neexistuje přímá korelace mezi počáteční délkou telomer a délkou života, nicméně některé syndromy se znaky předčasného stárnutí jsou s délkou telomer spojovány.
Kmenové buňky jsou nedifencované buňky schopné sebeobnovování a diferenciace na jiné buněčné typy.
Typy kmenových buněk:
Kmenové buňky proliferují a dávají vzniknout jednomu nebo více typům buněk
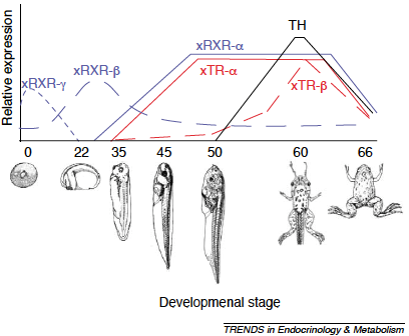
Metamorfóza řízena hormonem štítné žlázy – thyroxinem a jeho derivátem trijodthyroninem. T3 vzniká z thyroxinu působením enzymu dejodinázy typu II, působením dejodinázy typu III vzniká neaktivní derivát. T4 i T3 se s různou afinitou váží na jaderný thyroidní receptor (TR). TR vytváří komplex s retinoidním receptorem RXR a společně aktivují transkripci cílových genů.
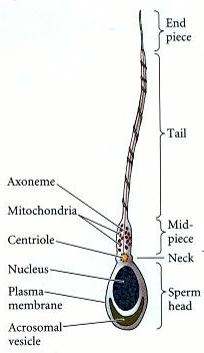
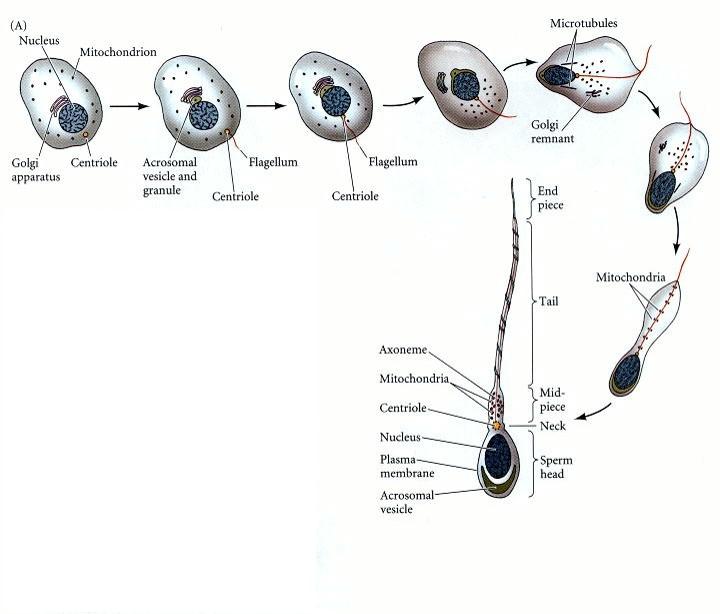
Spermie je samčí pohlavní buňka. Je pohyblivá. Do vajíčka přináší genetickou informaci a centriolu.
Skládá se ze tří částí:
V hlavičce se nachází genetická informace. DNA je v normální buňce navinutá na histony. Ve spermiích jsou však použité jiné bílkoviny, protaminy, které umožňují větší kondenzaci DNA. Součástí hlavičky je též akrozomální váček, který obsahuje enzymy potřebné pro porušení membrány vajíčka.
Krček obsahuje mnoho mitochondrií, aby měla spermie dost energie k pohybu.
Bičík se skládá z mikrotubulů. Pohyb bičíků je zajištěn dyneinem, což je molekulární motor.
Sexem se myslí vlastně vznik pohlavních buněk, jejich migrace k sobě (spermie k vajíčku „atrakce“, a jejich splynutí).
Reprodukce je vznik nového života, spojením dvou haploidních genomů vznikne nový život, který už je diploidní a vznikl kombinací dvou gamet
nepravidelná pigmentace kůže, narušené nebo chybějící nehty, později selhávání orgánů (kostní dřeň), plicní fibróza, predispozice ke vzniku nádorů
Tkáňové kmenové buňky se již v léčbě používají při transplantacích kostní dřeně a při kožních náhradách. Jsou však schopné diferenciace jen do omezeného počtu buněčných typů.
Při transplantanci kostní dřeně hematopoetické buňky dárce postupně osídlí tkáň příjemce a obnoví krvetvorbu. V určitých situacích je možné využít pupečníkovou krev jako zdroj kmenových buněk krvetvorby.
Kožní náhrady se využívají při léčbě popálenin. Kultivují se kmenové buňky keratinocytů. Proliferují a tvoří pláty pokožky, které jsou transplatované na poraněné místo. Nejedná se o plnohodnotnou kůži – chybí mazové žlázy a ochlupení.
Neoblasty jsou jediné mitoticky se dělící buňky v těle ploštenky. Za fyziologických podmínek zajišťují přirozenou obnovu buněk. Jsou schopné diferenciovat do všech buněčných typů (cca 40), které se vyskytují v těle ploštenky, včetně pohlavních buněk. Jedná se o kmenové buňky.
Průměrná délka života je charakteristika populace. Může se lišit mezi jednotlivými populacemi daného druhu.
iPSC
Indukované kmenové buňky jsou dospělé somatické buňky, které jsou uměle reprogramovány do stavu podobnému embryonálním kmenovým buňkám. Stačí k tomu kombinace čtyř transkripčních regulátorů. In vitro se diferenciují na požadovaný buněčný typ. Výhodou je, že pacientovi mohou být odebrány a indukovaný jeho vlastní buňky. Problémy jsou ve způsobů dodání reprogramujících faktorů a taky v tom, že množící se buňky jsou náchylné ke tvorbě nádorů.
Unipotentní buňky jsou schopné diferencovat na jeden buněčňý typ.
Př. epiteliální
Ploštěnka patří mezi prvoústé, kteří zatím nemají vytvořenou pravou tělní dutinu (mají schizocoel). Tělo tvoří tři zárodečné listy.
Tělo je dorzo-ventrálně zploštělé, což dalo jméno celé skupině. Má vysokou regenerační schopnost, která je známá po staletí.
V závislosti na dostupných zdrojích je schopná zvyšovat nebo snižovat počet tělních buněk. Nezáleží na rovině amputačního řezu. Pokaždé z fragmentů doroste nový jedinec (pokud obsahuje neoblasty). Gradienty morfogenů představují poziční informaci. Za určitých podmínek je schopná zregenerovat i z jediné buňky.
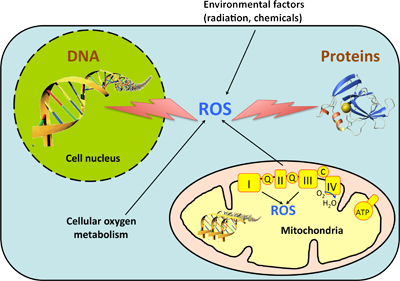
Frekvence vzniku mutací v mitochondriálním genomu je výrazně vyšší než v jaderném genomu (asi 10krát-20krát). V mitochondriích jsou méně účinné opravné mechanismy. Poškození mitochondrií může vést k poruchám produkce energie, nadměrné produkci reaktivních derivátů kyslíku a k indukci apoptózy.
Dochází k četným morfologickým změnám:
| pulec | žába | |
|---|---|---|
| Pohyb | svalnatý ocas s ploutví | ztráta ocasu, pohyb po čtyřech končetinách |
| Dýchání | žábry, kůže, larvální hemoglobin | plíce, kůže, dospělý hemoglobin |
| Potrava | herbivorní | karnivorní |
| Vylučování | amonotelní | ureotelní |
| Povrch těla | tenká epidermis, tenká vrstva škáry, chybí kožní žlázy | silnější epidermis i škára s dobřevyvinutými kožními žlázami |
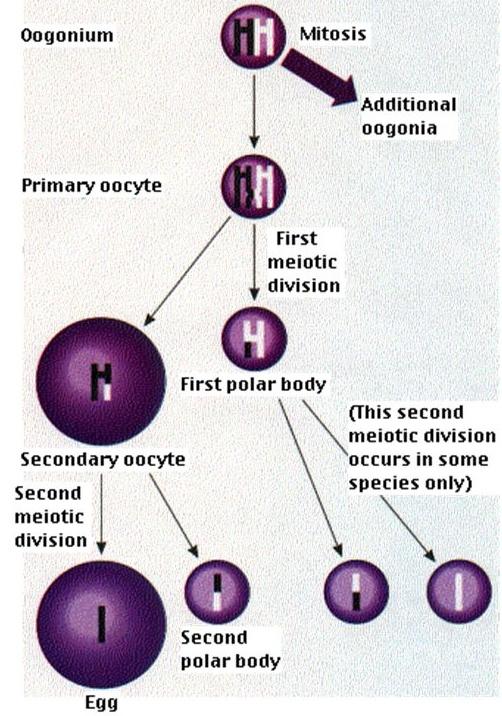
Oogeneze je proces při kterém se vytváří vajíčka. Jelikož vajíčko přináší cytoplasmu včetně zásobních látek do budoucího zárodku, je třeba příliš neplýtvat. Z toho důvodu se oogenezí vytváří z jednoho oogonia pouze jedno vajíčko.
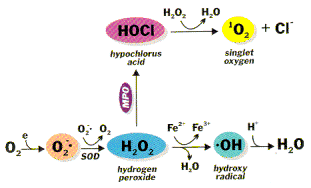
ROS (reactive oxygen species) jsou reaktivní formy kyslíku, které poškozují DNA, lipidy i proteiny. K jejich produkci dochází v mitochondriích. Asi 2-3 % kyslíku v mitochondriích jsou redukovány na ROS. Odstraňují se pomocí superoxid dismutázy (superoxidový radikál → H2O2), katalázy (H2O2 → H2O), glutathion peroxidázy.
Neexistuje vždy korelace mezi rychlostí metabolismu a délkou života (rychlejší metabolismus…– kratší život). Existuje celá řada výjimek jako ptáci, vačnatci, netopýří… Ač je tělo rypoše vystaveno velké míře oxidativního stresu, dožívá se v zajetí 28 let. Tedy musí mít jiné mechanismy.
Př.: peroxidy, superoxidy, hydroxylový radikál, singletový kyslík.
Hutchinsonův-Gilfordův syndrom
Lidé s progerií mají mutaci v genu kódujícím lamin A, což je složka jaderné laminy. Postižení jedinci se dožívají v průměru 12 let. Vykazují mnoho znaků spojených se stárnutím.
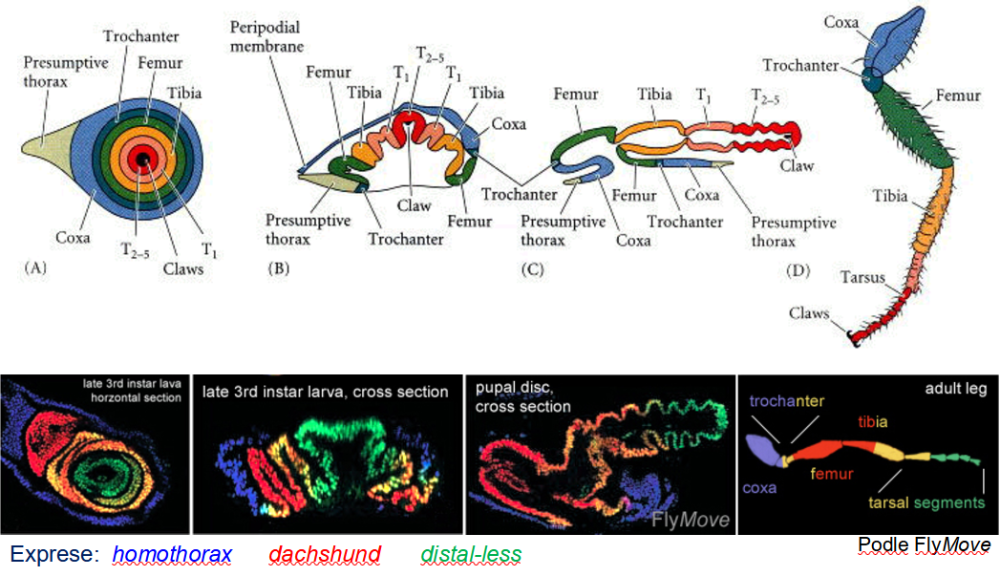
Imaginální terčíky jsou skupiny buněk, které si během embryonálního vývoje zachovají nediferenciovaný charakter. Během larválního vývoje proliferují víceméně nezávisle na okolní larvální tkáni.
Před metamorfózou struktura „složeného teleskopu“.
Během metamorfózy dochází k elongaci, „vysunutí teleskopu“, a specifikaci jednotlivých částí daného orgánu – nohy, tykadla, křídla…
Multipotentní buňky jsou schopné diferenciovat na více než jeden buněčný typ určité tkáně (např. neurony, astrocyty, oligodendrocyty z neurálních kmenových buněk).
Př. tkáňové – neurálním střevní, hematopoetické…
Při neotenii se zachovávají juvenilní znaky u pohlavně dospělých jedinců (soma se ve vývoji zpožďuje za zárodečnou lini / pohlavními orgány – příklad heterochronie). Typický výskyt u axolotla (rod Ambystoma). Může být pernamentní nebo fakultativní (v závislosti na podmínkách prostředí).
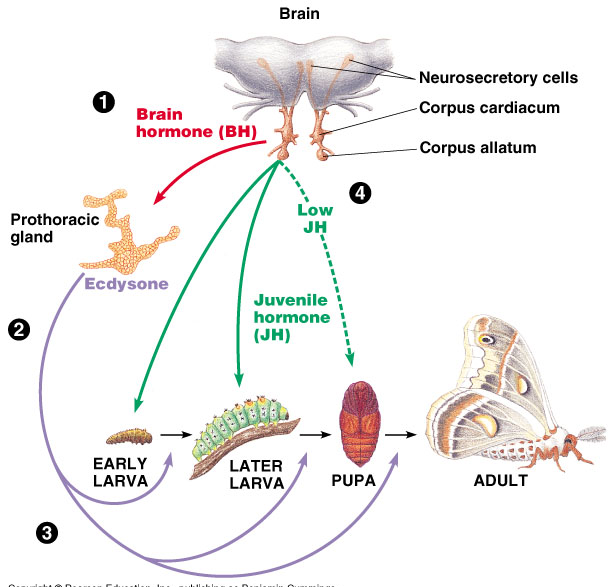
prothoracicotropic hormone, brain hormone
PTTH stimuluje prothorakální žlázu k produkci ekdyzonu.
S oogenezí je spojena folikulogeneze, vznik, vývoj folikulu: oocyt se nevyvíjí osamocen, je obalen buňkami, je součástí folikulu, ten je vyplněn folikulární tekutinou (antrum).
Jeho růst je závislý na hladině:
K oplození dochází zhruba v horní třetině vejcovodu.
Vajíčko se neuvolňuje samo, ale s kumulárními buňkami, tvoří cumulus oophurus – vejconosný hrbolek (těmi je přidělané předtím k folikulu zevnitř). Společně se uvolní do nálevky vejcovodu.
Maximální délka života je nejvyšší známý věk, kterého se organismus může dožít. Je charakteristická pro daný biologický druh.
#otazka Co jsou extraembryonální tkáně? Jedním z příkladů je ? žloutkový váček?
Totipotentní buňky jsou schopné tvořit celý organismus, včetně extraembryonálních tkání.
Př. zygota.
Pluripotentní buňky jsou schopné tvořit buňky všech tří zárodečných listů včetně buněk zárodečné linie. Netvoří buňky extraembryonální tkáně.
Př. embryonální kmenové buňky (ESC).
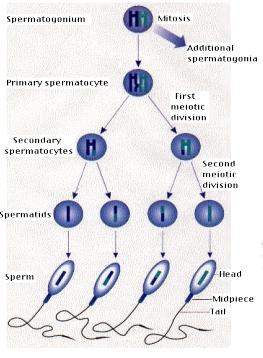
Výsledný počet buněk. Základní rozdíl je v tom, že z jedné diploidní spermatogonie (mitoticky se dělící buňka) vznikají ve dvou po sobě jdoucích děleních meiózou spermatidy (4), které jsou haploidní. V oogenezi vzniká pouze jedno haploidní vajíčko.
Čas, po který probíhá.Spermatogeneze probíhá po celý život, oogeneze jen v časném zárodečném vývoji.
Spermatogeneze je proces, během kterého se vytváří spermie. Z jednoho spermatogonia vzniknou čtyři spermie.
Juvenilní hormon udržuje organismus v larválním stádiu. Kontroluje expresi genů potřebných pro larvální vývoj. Je produkován v corpora allata. Patří mezi seskviterpeny. U jednotlivých skupin hmyzu se struktura může mírně lišit (např. u motýlů). Analogy JH (tzv. juvenoidy) se používají jako insekticidy.
Omezení příjmu potravy je jedním z mála zásahů, které prokazatelně mohou prodloužit věk, snižuje úroveň oxidativního poškození.
spermiogeneze
Spermatolyoza je proces, během kterého se buňka spermatidy přestavuje do podoby spermie.
Ekdyson neboli svlékací hormon řídí svlékání a metamorfózu. Indukuje expresi genů potřebných během svlékání a metamorfózy.
Váže se na ekdyzonový receptor (EcR – podobný receptoru pro thyroidní hormon). EcR tvoří dimer s proteinem Ultraspiracle (Usp, homolog RXR), EcR-Usp komplex indukuje expresi genů. V odpovědi na zvýšenou hladinu ekdyzonu je možné na polyténních chromozomech sledovat tvorbu „puffs“, které představují oblasti aktivní transkripce.
Je produkován prothorakální žlázou. V periferní tkáni je modifikován na aktivní formu – 20-hydroxyekdyson. Patří mezi steroidní hormony.
Sirtuiny jsou deacylázy (hlavně deacetylázi), mění postranskripční modifikace velkého množství proteinů (transkripční faktory, enzymy – SOD, komplex I, II…).