Created by Kamila Klavíková
Zápisky z přednášky MB170P75 na PřF UK s Lindou Nedbalovou a Tomášem Herbenem.
Podobní teplokrevní obratlovci se se snižující teplotou zvětšují. Takový jedinci mají lepší poměr povrch : objem, čímž se zmenšuje povrch, kterým se může odpařit teplo.
Organismy se stejným objemem těla mohou mít různě velký tělní povrch. Směrem do chladných podmínek se velikost tělních externit zmenšuje, což pomáhá snížit tělní ztráty.
Psychrofilní organismy = „chladnomilné organismy“
Poškození chladem – způsobuje změny v semipermeabilitě a fluiditě membrán, což vede k úniku důležitých iontů.
Poškození mrazem – led se začíná tvořit okolo kondenzačních jader, což má za následek mrznutí čisté vody a růst koncentrace rozpuštěných látek v roztocích.
Vymrzání extracelulární vody způsobuje odčerpávání vody z buněk ven, což vede ke zvýšení koncentrace rozpuštěných látek. To má za následek, že obsah buněk hůře mrzne.
Kryptoreaktanty jsou látky, které zabraňují vzniku intracelulárního a extracelulárního ledu. Př. vyšší koncentrace NaCl, specifické peptidy, glykoproteiny.
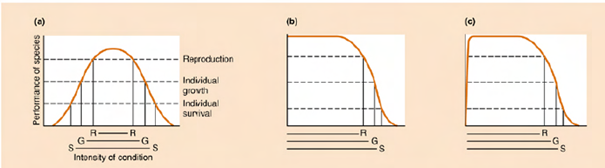
Častěji než letální rozhodují suboptimální podmínky. Globální klima výrazně koreluje s rozšířením řady druhů, ale závisí na mikroklimatu.
Je to souhra teploty a dalších faktorů:
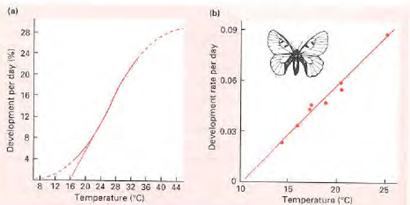
Vliv teploty na rychlost vývoje růstu je lineární – day-degrees
Aklimatizace probíhá znatelně hůře než aklimatizace na nízké teploty. Dochází k denaturaci proteinů, inaktivaci enzymů a dehydrataci. Organismy jsou schopné přežívat díky dehydratovaným stádiím – spory, cysty, semena.
Př. teplomilných organismů: Alvinella Pompejana, Thermus aquaticus
Endotermní organismy – „termostat“ v mozku
Všechny organismy mají tendenci ztrácet vodu (vylučování, ochlazování). Interakce s rychlostí větru. Řada organismů vyžaduje téměř 100% relativní vlhkost.
Salinita ovlivňuje počet orgismů v místě ostrých předělů mezi mořskými a sladkovodními biotopy. Vysoký osmotický tlak v původním roztoku. Dva typy rostlin v reakci na salinitu: obligátní halofyty (vyžadují) a fakultativní halofyty (snáší).
V hlubokých jezerech a na dně oceánů jsou extrémní tlaky. Mají přímý i nepřímý vliv na organismy. Rostoucí tlak zvyšuje rozpustnost CaCO3, což vede k redukci koster u hlubinných organismů.
Podle citlivosti na kolísání tlaku rozlišujeme stenobatické druhy (citlivé) a eurybatické druhy (tolerantní).
Vizkozita vody je výrazně ovlivněna teplotou. Má velký význam pro sedimentaci nepohyblivých organismů.
Hustota vody se mění v závislosti na teplotě – vzniká tzv. „hustotní anomálie vody“. U vodních organismů odpadá mechanická limitace, proto mohou dosahovat velkých rozměrů.
Různé organismy během evoluce vyvinuly různá opatření, jak se sám nestát potravním zdrojem.
Rostliny mají
Různé části rostliny jsou chráněné do různé míry.
Živočichové mají adaptace
Atmosférické proudění ovlivňuje šíření organismů.
Přímý toxický efekt: koncentrace H+ a OH− – < 3 a > 9 → destrukce protoplasmy buněk hodnot. Dochází k porušení osmoregulace a ovlivnění aktivity enzymů.
Nepřímý efekt pH ovlivňuje dostupnost a přítomnost dalších iontů. Dochází ke zvýšení koncentrace toxických iontů těžkých kovů. Působí na kvalitu zdrojů nutných pro organismus. Př.: < 4,0-4,5 – toxický Al3+
Stenobatické – citlivé druhy ke kolísání tlaku
Eurybatické – tolerantní ke kolísání tlaku
Aklimatizace je přizpůsobování se novým žívotním podmínkám.
Úspěšnost jedince je silně závislá na intenzitě faktorů (podmínek) jako je vlhkost, atmosferický tlak, hydrostatický tlak, proudění vody...
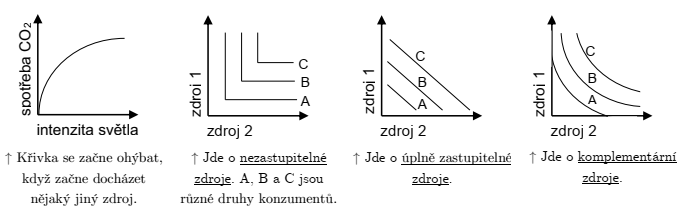
Pro zdroje platí, že aktivitou organismu jejich dostupnost klesá.
| Podmínky | Zdroje |
Teplota | Záření |
Přítomnost vody | CO2 |
Atmosférický tlak | Minerální látky |
Proudění | Organismy – potrava |
Salinita | Prostor |
Hydrostatický tlak |
|
pH |
Limitujícím zdrojem je ten, jehož koncentrace určuje populační růst.
Jinými slovy, pokud mám několik nutných „ingrediencí“, omezuje mě hladina toho zdroje, kterého mám nejméně. Naopak se nemusím tolik zajímat o hladinu zdrojů, u nichž mám relativní nadbytek. Ekologové toto nazývají Liebigův zákon minima.
Fundamentální nika – nika, kterou by organismus zaujal, pokud by nebyl omezovaný jinými organismy.
Realizovaná nika – nika, kterou organismus reálně v daném ekosystému zaujme.
Zdroje jsou vše, co organismus využívá a co potenciálně ovlivňuje jeho fitness a v důsledku toho populační růst (v ekologii zásadní).
Zdroje jsou spotřebovatelné a důležité k růstu jedinců a populací. Aktivitou organismu množství zdroje klesá.
Mohou být jak potravní, tak nepotravní. Mezi nepotravní zdroje patří např. hnízdiště a místa k přenocování.
| Teplota, pH | Jedy | Minerální látky |
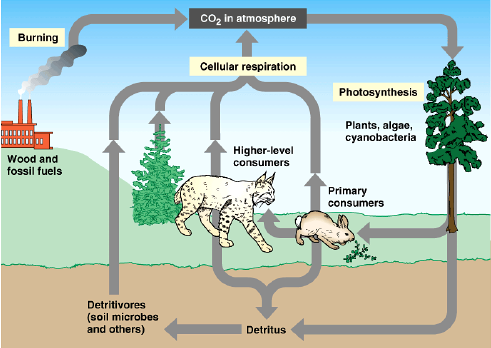
Živočichové a houby jsou, na rozdíl od rostlin, heterotrofové což znamená, že si nemohou sami syntetizovat uhlíkové cytoskelety z anorganických sloučenin uhlíku. Proto musí požírat uhlík obsažený v organických látkách.
Rozdělme je na rozkladače (požírající DOM—dead organic matter) a predátory (v širokém slova smyslu, tedy konzumenty rostlinné či živočišné potravy).
Pro herbivory nastává problém v tom, že z rostlin získavají výrazně větší podíl uhlíku. Navíc lignin (abundantní aromatický polymer v rostlinách) je téměř vždy nestravitelný. Obecně jsou pro herbivory atraktivní plody, semena a roustoucí pletiva, protože obsahují více aktivních enzymů.
NPP/plocha nejvyšší v tropickém lese a na korálových útesech.
NPPL/plocha volného oceánu je nízká, ale vzhledem k velké ploše NPP srovnatelná s tropickými deštnými lesy. V oceánu je však o tři řády nižší biomasa (absence podpůrných a vodivých pletiv).
Hlavní podíl (75 %) na globálních antropogenních emisích CO2 do atmosféry má spalování fosilních paliv. Dalším významným zdrojem je změna ve využívání krajiny.
Pro čerpání vody je maximalizován objem kořenů. Uplatňuje se mykorhiza: mutualistické soužití kořenů vyšších rostlin s houbami, které dodávají minerální živiny rozpuštěné ve vodě.
Mezi minerální živiny patří: N, P, S, Ca, K, Mg... Většinou se vyskytují jako kationty (jako anionty se vyskytují v půdě např. nitráty, ty jsou méně rozpustné). Výskyty N a P bývají limitující.
Dusík se vyskytuje v několika formách. Mezi anorg. formy patří NO3 a NH4 v půdě a N2 ve vzduchu. V organických sloučeninách se vyskytují typicky jako redukovaná skupina NH2+, a to například v mrtvé hmotě a uvnitř těl (v aminokyselinách a dusíkatých heterocyklech).
Dostupnost fosforu závisí na jeho přítomnosti v geologickém substrátu a transportu větrem. V půdě se nacházejí látky s fosfátovými skupinami.
N je přitahován do kořenů mnohem rychleji než P, který je naopak v půdě pevně vázán (hustý kořenový systém výhodou).
Důležitým zdrojem je záření (tedy tok radiační energie), jednodušeji řečeno světlo, respektive fotosynteticky aktivní část záření (PAR, také FAR). Účinnost využití PAR je vlastně velmi malá... Využitelnost záření závisí na vodě. Dilema více vody versus rychlejší fotosyntéza.
Konečně oxid uhličitý je samozřejmě zásadním zdrojem pro fotosyntetickou aktivitu.
U rostlin existuje poměr investice v kořenovou část vůči investice v část nadzemní, nazýváme jej root:shoot ratio. Rostlinu investuje více do toho nedostatkového, tedy v případě toho, že je chudá půda, více růstu bude probíhat v podzemí, jelikož právě živiny v půdě jsou limitujícím faktorem.
U fotoautotrofů obecně zkoumáme míru probíhající fotosyntézy v porovnání s mírou respirace. Stav, kdy se rychlosti obou procesů rovnají, označujeme jako kompenzační bod.
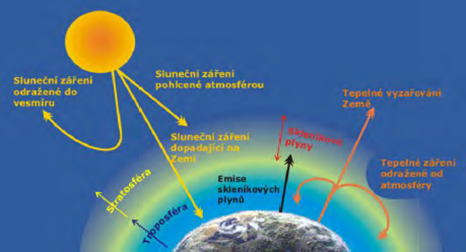
Sopečná činnost nejvíce produkuje H2O, CO2, SO2. Mezi další látky patří H2S, H2, CO, HCL, HF, He.
Podíl vulkanické činnosti na produkci skleníkových plynů je kompenzován prostřednictvím ochlazování daného zastíněním sopečným popelem a prachem („haze effect“).
Mezi skleníkové plyny patří vodní pára (60 %), CO2 (cca 26 %), CH4, N2O, O3.
Bez výskytu skleníkových plynů by na zemi bylo 18 °C.
Nutriční hodnota těl rostlin a živočichů, která heterotrofové požírají, se odvíjí od poměru uhlíku a dusíku.
Živočichové mají v těle hodně bílkovin, jež ve své struktuře obsahují právě dusík—rovnováha je tedy vychýlena ve prospěch dusíku.
Zato u rostlin, s vysokým obsahem polysacharidů, je poměr posunut ve prospěch uhlíku.
Složení živočišných těl kolísá je stabilnější než u rostlin. V praxi nastávají dysbalance, které vedou například k tomu, že mšice vylučuje nadbytečný cukr z rostlinového floému ve formě sladké šťávy.
Primární produkce je dána fotosyntetickou činností organismů a jejím produktem je biomasa.
Organismy pomocí fotosyntetického fotoaparátu využívají jako energii sluneční záření a zdrojem uhlíku je pro ně CO2. Vytvořená organická hmota tvoří základnu pro další produkci, a tedy i výskyt organismů.
Producenti nejen že vytvářejí organickou hmotu, ale částečně jí využívají na vlastní metabolismus. Veškerá vytvořená organická hmota vytvořená za časové období je hrubou primární produkcí (brutto). Hrubá produkce zmenšená o vlastní metabolickou spotřebu organismu. Označuje se jako čistá primární produkce (netto).
Hrubá (GPP) - energie (uhlík) vázaná/ý na fotosyntézu za jednotku času = intenzita fotosyntézy.Je to veškerá vytvořená organická hmota vytvořená za časové období. Množství uhlíku fixovaného fotosyntézou.
Čistá (NPP) = hrubá – respirace – hrubá produkce zmenšená o vlastní metabolickou spotřebu.
Respirace: energie vynaložená na metabolismus, ztráta uhlíku dýcháním rostlinou: NPP = GPP – R
Kompenzační bod: fotosyntéza = respirace (veškerá produkce jde na udržovací metabolismus)
Okamžitá biomasa: biomasa v okamžiku pozorování/sklizně
Použití radioizotopů 14C – přidání známého podílu
Solární konstanta = 8,37 J x cm-2 x min -1
Fotosynteticky aktivní záření (390-710 nm)
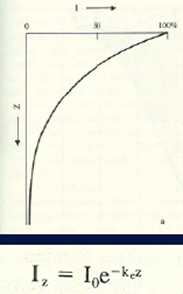
Záření ve vodním prostředí
Schopnost využívat zdroje je základní ekologickou charakteristikou populace. Úspěšnější je ta populace, která dokáže konzumovat, tedy existovat při nižší hladině zdroje: má nižší úroveň kompenzačního bodu.
Kompenzační bod R* značí hladinu zdroje, při které je populační růst nulový.
Koloběh uhlíku má úzkou vazbu k toku energie. Převážně plynný cyklus poháněný protichůdnými silami fotosyntézy a respirace.
Roste role litosférického zásobníku v důsledku spalování fosilních paliv.
Roste konc. CO2 v důsledku spalování fosilních paliv a kácení trop. pralesů.
Zásoby (pools) chemických prvků jsou uloženy v různých rezervoárech:
Biogeochemie se zabývá studiem cyklů chemických prvků UVNITŘ a MEZI zásobníky
Biogeochemické cykly – rozlišení podle typu základního rezervoáru:
Na rozdíl od energie mohou být chemické látky použity opakovaně a jejich cyklování je výrazným rysem – klíčová role rozkladačů
Výrazný vliv organismů včetně člověka
6CO2 + 12 H2O à C6H12O6+ 6O2 + 6 H2O
Bilance CO2 – změna množství v okolním vzduchu za čas
Bilance O2 – výhodná ve vodním prostředí – princip tmavé a světlé lahve à změří se obsah kyslíku v lahvi na počátku a voda se dá do průsvitné a černé lahve a nechá se fotosyntetizovat à na konci se změří obsah kyslíku
V koloběhu síry je významná atmosférická i litosférická fáze.
Síra se do atmosféry uvolňuje mořským aerosolem, vulkanickou činností a anaerobně. Zpětný tok z atmosféry oxidací na síranové ionty.
Do vody se dostává zvětráváním hornin a z atmosféry.
Do potravních řetězců se dostane malý podíl.
Ztráty do sedimentů → tvorba Fe2S
Hlavním zdrojem vody jsou oceány. Podíl vody, která se aktuálně přemisťuje, je malý (asi 0,08 %). Hraje však klíčovou roli pro suchozemská společenstva. Cyklus by probíhal i bez přítomnosti organismů, ale vegetace hraje významnou roli.
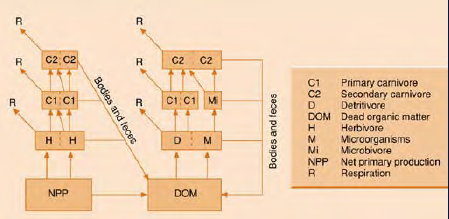
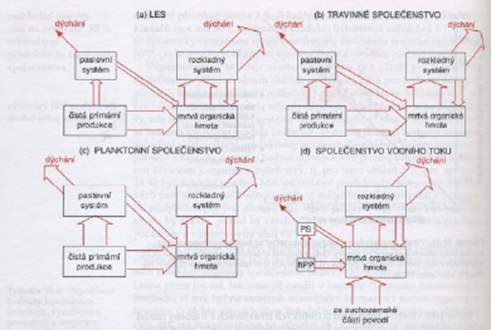
Sekundární produkce je rychlost produkce nové biomasy heterotrofními organismy.
Bakterie, houby a živočichové svou hmotu odvozují od NPP přímo nebo nepřímo. Mezi sekundární a primární produktivitou obecně platí pozitivní vztah.
Potravní řetězce – pyramida pastevního systému, dekompozice
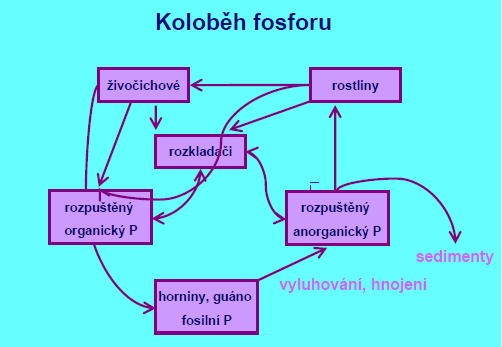
Sedimentární typ – vždy opouští pevninu a včleňuje se do sedimentů. Hlavními zásobníky fosforu jsou mořské sedimenty a horniny, voda.
Koloběh fosforu je ovlivněný činností člověka – zemědělství, lov mořských ryb, čistící prostředky – eutrofizace.
Uvolnění P zpět do vodního sloupce na rozhraní sediment-voda záleží na redox potenciálu povrchové vrstvy bahna – ten určuje formu iontů Fe: ox. podm. – nerozpustný FePO3, re. podm. – rozpustný Fe3 (PO3)2.
Zdrojem P je zvětrávání substrátu, hlavní zásoby jsou v sedimentech, o uvolňování ze sedimentů rozhoduje koncentrace rozpuštěného kyslíku ve vrstvě nade dnem.
Formy: DP (SRP), PP, pro primární producenty dostupný jen SRP
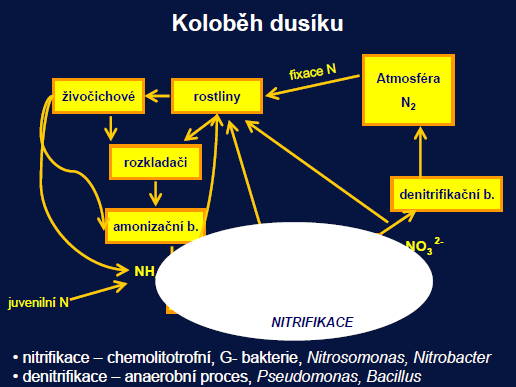
Plynný cyklus – nejvýznamnější atmosférická fáze. Atmosférická fixace (výboje blesků) 4 %, zbytek biologická fixace – nutná energie k rozštěpení trojné vazby.
Ovlivnění činností člověka – odlesnění (nárůst dusičnanů ve vodách), hnojiva – eutrofizace, spalování fosilních paliv a zemědělství – oxidy dusíku – acidifikace
Průmyslová fixace stejná jako přirozená
Pravděpodobně silně podceněná role litosférického zásobníku
Min. 3–5 % sušiny buněk
Formy: DN (NO3-, NO2, NH4+), PN, TN, pro organismy energeticky nejvýhodnější NH4+
Poměr NPP produkcí trofických hladin se nazývá Lindemanova účinnost.
r stratégové upřednostňují reprodukční rychlost a mobilitu, zato kvalita a konkurenceschopnost pokulhává.
Typický r-stratég je organismem obsazujícím narušená stanoviště – jedná se o primární fázi sukcese.
Písmeno r je matematický symbol pro rychlost reprodukce (rate of reproduction).
K stratégové jedou na kvalitu. Prostě opak r. Typický K-stratég bude mít málo velkých mláďat (resp. semen), o které se bude starat. Příkladem může být slon, člověk nebo dub.
Během sukcese K-stratégové obvykle významněji nastupují až v pozdních fázích a postupně vytlačují r-stratégy.
Písmeno K je matematický symbol pro únosnou kapacitu prostředí, tj. maximální podíl přeživších jedinců.
Volně žijící Azobacter, symbiotické Rhizobium
Klíčová role bakterií.
Biologická fixace dusíku – heterotrofní bakterie z řady skupin (nejznámější hlízkovité b. bobovitých rostlin – Rhizobacter), sinice – heterocyty
Disperze je prostorové uspořádání jedinců. Pojem označuje i způsob, kterým se organismy rozšiřují.
Výjimečně zůstavají potomci u svých rodičů, zpravidla se uchycují jinde. U většiny organismů je pohyb každodenní součástí života. Přemisťování organismů vede k rozmanitému rozmístění jedinců v prostoru.
Záleží z jaké perspektivy se na to díváme.
Př. výskyt mšic: 1) z hlediska lesa → agregované, 2) z hlediska větve → náhodné, 3) z hlediska listu → pravidelné rozmístění).
Agregovaná – org. jsou se shlucích (V/M ˃ 1)
Náhodná – rozmístění se neodvíjí od vzájemných interakcí (V/M ≈ 1)
Rovnoměrná – plus mínus stejné vzdálenosti (V/M ˂ 1)
Obvykle se jedná o masový přesun organismů. Označuje jak transkontinentální přelety sarančat, tak i každodenní přesun krabů reagujících na příliv vody.
Téměř vždy se jedná o přesun z míst s nedostatkem potravy do míst s hojností potravy (sezónní výskyt druhů). Motivace k migraci může být i aktuální hustota jedinců.
Nosná kapacita prostředí je bod K, kde se protíná křivka natality a mortality. Natalita kompenzuje mortalitu. Protože na ní působí mnoho vlivů, spíše interval.
př. nezmar, korálové útesy – zooxantelly
Řasy v tkáních různých organismů
Lišejníky = mutualismus řas a hub
Rostliny jsou „lokální sebevrazi“, fixují dusík, čímž zlepšují podmínky pro jiné rostliny, se kterými prohrávají kompetici.
Bakterie – stimulace k zapnutí genů pro tvorbu hlíz → vytvoření hlízy → cévními svazky jsou dopravovány asimiláty z fotosyntézy a odváděny dusíkaté látky
Většina vyšších rostlin nemá kořeny, ale mykorhizu. Mykorhiza je těsné spojení rostlinných tkání a pletiv hub.
hlavně stromy
hyfy dovnitř buněk kořenů
Označení skupiny jedinců stejného druhu (mohou si vyměňovat genetickou informaci). Pojem se liší druh od druhu. Hranice populace mohou být určeny jednoznačně (jeleni v oboře) i uměle (mšice na listu, mšice na stromě, mšice v lese).
Vývoj natality a mortality se v závislosti na hustotě mění – natalita klesá, mortalita stoupá (přepočteno na jedince). Rozdíl mezi křivkami na grafu je čistý přírůstek.
Zaneseme-li vývoj mortality a natality do jednoho grafu, křivky se nám vždy protnou v bodě K. Bod K se nazývá nosná kapacita prostředí. Natalita kompenzuje mortalitu. Protože je populace ovlivněna mnoha vlivy, je K spíše interval než bod.
denzita ˂ K → natalita vyšší, populace roste
denzita ˃ K → mortalita vyšší, populace klesá
mortalita = natalita → početnost se nemění, hustota, ke které má populace konvergovat (směřovat) (neuvažujeme imi/emigraci)
Sezónní cykly jsou výrazné u některých organismů, kdy část sezóny převažuje mortalita a část natalita (např. mnoho semenáčků umírá hned po vyklíčení).
ln λ se značí jako r = vnitřní rychlost růstu, kde λ představuje zákl. čistou reprodukční rychlost.. Vyjadřuje rychlost, za jakou se změní populace za jednotku času. r˂0 – populace vymírá; r˃0 – populace roste
Organismy v trávicích traktech (extracelulární symbionti) = pevná vazba mezi mikroorganismem a tělem partnera. Tráví celulózu, syntetizují vitaminy. Mikroorganismy – neustálý příjem potravy, stabilní podmínky.
Alogenní sukcese je sukcese vyvolaná vnějšími geofyzikálními vlivy. Je to v zásadě věc botanická, protože rostliny tvoří nejvíce biomasy a nejvíce přispívají do nekromasy. Živočichové spíše pasivně odrážejí rostlinou sukcesi (ale není tomu tak vždy).
Definice založeny na reprodukčních barierách, geneticky i ekologicky.
Ekologická definice. Druh je tvořen spřízněnými jedinci mezi nimiž je kompetice vyšší než vůči jedincům jiného druhu.
Každý druh si hlídá svou niku = hybridi nemůžou konkurovat čistým jedincům.
Cílem je udržet minimální mezidruhovou a vysokou vnitrodruhovou kompetici.
K primární sukcesi dochází na panenské půdě, kterou ještě neovlivňovalo žádné společensvo.
Př.: nové písečné přesypy, lávový proud, nově vzniklý sopečný ostrov
Př. Na pobřeží jezera Michigan je rozsáhlá sekvence písečných dun. Na pohyblivých hřebenech už je nějaká tráva. Po sto letech je nahrazena jalovci a zase nějakou jinou trávou. Za 150 let by tu už mohli být jehličnany a do 400 smíšený les.
Denzita je počet jedinců na jednotku.
Průměrná denzita je podíl počtu jedinců a plochy daného habitatu. Je velmi choulostivá na náš úhel pohledu.
Př. denzita mšic: 1) z hlediska lesů a polí je velmi malá, 2) z hlediska hostitelského stromu je docela vysoká).
Př. v USA žije 75 lidí na km2, hodně území je ale téměř neobydleno, a města jsou přecpaná → kolem průměrného Američana je 3630 lidí (velký rozdíl než minulá hodnota 75, může mít za následek třeba migraci).
„Normální“ denzita je denzita vztažená na jednotku zdroje.
Efektivní denzita je vážený průměr.
Př. na 10 kytkách po 91 housenkách, na 90 kytkách po 1 housence (10×91 + 90×1 = 1000 housenek) → 910 housenek má denzitu 91 a 90 housenek denzitu 1 → (910×91 + 90×1)/1000 = 82,9 housenek na kytku
Základní reprodukční rychlost (R0) je celkový počet potomků dělený počátečním počtem jedinců. Udává kolikrát se zvětší populace za generaci v diskrétně se rozmnožující populaci.
Základní čistá reprodukční rychlost (λ, protože chceme rychlost ne v generacích ale ve standardních časových jednotkách) → kolikrát se zvětší (zmenší) populace za jednotku času. Je zde zakombinováno jak přežití rodiče, tak vznik nového jedince.
Př. λ=2 → rodič zemřel a má 2 potomky, nebo rodič přežil a má 1 potomka
Mutualismus je oboustranně výhodná interakce mezi organismy.
Mutualisté tvoří většinu biomasy.
K autogenní sukcesi dochází na nově vytvořených stanovištích, bez vnějších abiotických podmínek.
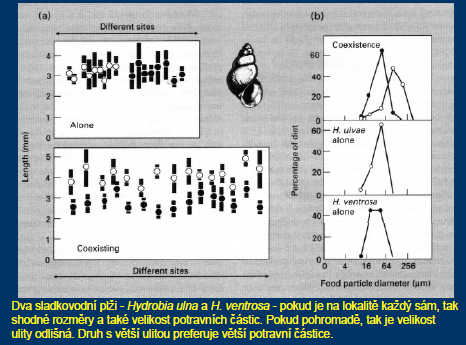
Pokud koexistují dva skutečně si konkurující druhy – a to ve stabilním prostředí – pak je koexistence možná pouze v důsledku nikové diferenciace, tedy díky částečnému odlišení realizovaných nik konkurujících si druhů.
K sekundární sukcesi dochází na místech, kde došlo k částečnému nebo úplnému odstranění druhů, ale zůstala vyvinutá půda se semeny a sporami.
Př.: opuštěná zemědělská pole
Metapopulace je komplex subpopulací, na které se ta původní rozpadla (např. kvůli predaci).
Mohou se navzájem ovlivňovat emigrací, imigrací a osidlováním nových habitatů. U samotné subpopulace se může jevit jiný závěr (např. vymření), než když zkoumáme celou metapopulaci → navzájem se ovlivňuje
„Rozdíl tělesných hmotností sousedících druhů je roven přibližně dvojnásobku, zatímco délkové míry je liší relativně o 1,3x.“
Sukcese je nesezónní, směrovaný a kontinuální proces kolonizace a zániku populací jednotlivých druhů na určitém místě. Může mít různý časový rozsah. např. depozici ovčího bobku až k dekompozici nebo od poslední doby ledové do teď.
Degradační sukcese je sukcese společenstev na mrtvé organické hmotě.
ptáci, savci, hmyz...
Výsledná podoba a postup, jak jí dosáhnout, je předem „naprogramován“. Výsledná podoba je determinována a predikovatelná.
rostliny, korálovci, trubýši...
Rostou soustavným přirůstáním nebo oddělováním dalších částí (větve, oddenky, polypy). Mají neukončený vývoj.
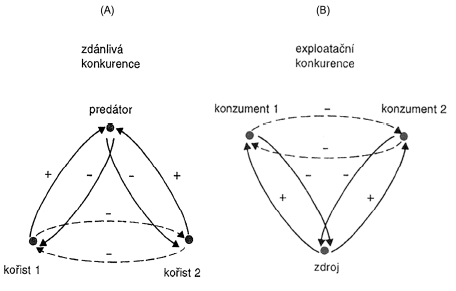
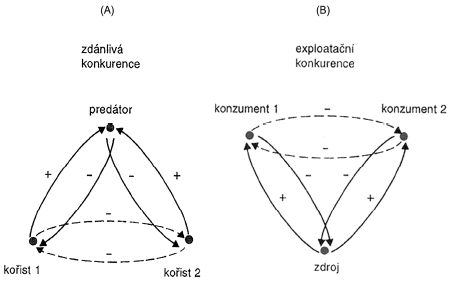
Zdánlivá konkurence – prostor bez predátora
Exploatační konkurence – nedochází k bezprostřednímu střetu mezi kompetitory
Interfernční konkurence – dochází k přímému mezidruhovému střetu
Klimax je koncepce konečného stádia sukcese.
Nejdříve myšlenka monoklimaxu, ale ta byla napadána, protože podoba konečného stádia bude záležet na mnoha lokálních faktorech (polyklimax). Stabilní klimax asi neexistuje – různá narušení, tzv. disturbance.
life tables, fecundity tables
Tabulky přežívání a plodnosti se často uvádí na základě počtu samic, protože se lépe přiřazuje narození mláďat. Problém je s populacemi s nepoměrem samců a samic (včely).
Statistickou tabulku přežívání využíváme v případě, kdy nejsme schopni rozdělit kohorty, ale můžeme určit věk. Je to věková struktura populace.
Organismy jsou nerozlučně pohramadě, v blízkém fyzickém vztahu.
 |  |
Disturbance je narušení ustáleného stavu, změna procesu. Je to jakákoli událost v čase, která odstraňuje organismy, nebo jinak mění společenstvo z hlediska dostupnosti prostoru, zdrojů nebo fyzikálních podmínek. Obvykle má za následek uvolnění prostoru nebo jiného zdroje, který je pak k dispozici jiným organismům.
Př. přírodní katastrofy, glaciace, globalní oteplování, mezidruhová kompetice...
Ranně sukcesní druhy jsou první kolonisté. Mohou změnit podmínky a tak usnadnit kolonizaci dalším druhům = facilitace, např. změnou vlastností půdy – hromadění N, změna hustoty a pH,...
Inhibice – opak facilitace, rychlý kolonista zabere pro sebe rozhodující zdroje.
Kořist není rozmístěna náhodně díky heterogenitě prostředí – predátoři se přemisťují mezi oblastmi, které se liší hustotou kořisti – agregrace v oblasti s vyšší hustotou.
Vznik částečných prostorových refugií – část populace kořisti bude mít menší pravděpodobnost predace.
Pro věkově specifické tabulky plodnosti potřebujeme znát i plodnost jednotlivých věkových tříd (každá má jinou fekunditu).
Kohortová tabulka přežívání udává, kolik jedinců se dožilo určitého věku.
Uvádíme věkové skupiny, počet přežívajících jedinců do roku x (ax), podíl přežívajících z původní kohorty k roku x (lx), počet narozených ♀ v roce (Fx), počet narozených ♀ připadajících na přežívající ♀ (mx), atd…
Predátor málokdy vyhubí jeden druh a pak se vrhne na další, užírá ze všech. Efekt masožravých predátorů – odstranění hvězdic ze skalních útesů – rozvoj kompetičně dominantních slávek a následný pokles druhů.
Predace je konzumace organismu jiným organismem.
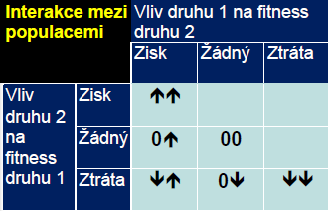
Účinky predace na kořist – negativní – snížení fitness, snížení početnosti
Útok veden proti nejslabší kořisti
Kompenzační reakce většinou výsledkem omezené vnitrodruhové kompetice – zejména u rostlin (zvýšení fotosyntézy, mobilizace zásobních cukrů, spící pupeny,…)
Searching + handling
Krátký handling – generalisti (zpěvní hmyzožraví ptáci)
Krátký searching – specializace (lvi, volavky,…)
maximalizace čistého zisku energie (handling time X searching time)
Polyfágní dravci – preference potravy a její změny
Monofág – pouze jedna živná rostlina
Oligofág – požírá více druhů, které jsou zpravidla příbuzné
Výsledkem jednoduchých modelů nestabilní kolísání a vymření dravce i kořisti, ale realita mnohem složitější.
Nesmíme zapomenout, že vztahy kompetitor-kompetitor nebo dravec-kořist, má vliv i na jiné organismy.
Nepřímé efekty tzv. keystones. Odstranění predátora nemůsí hned znamenat nárůst jeho kořisti, protože zde může být někdo „třetí“ a i další, koho to ovlivní a výsledek je najednou jiný
Top-down neboli kontrola shora. Konzumenti regulují počet zdrojů např. otázka Proč je svět zelený? Protože predátoři drží hebivory na uzdě a zeleň se hromadí podle ne/přítomnosti predátora se biomasa přelévá z jedné trofické hladiny do druhé = kaskádový efekt
Botton-up neboli kontrola zdola. Konzumenti jsou regulováni svými zdroji stejná otázka – Protože se některé rostliny naučili bránit a herbivoři kompetují o malé množství nechráněných rostlin a zároveň predátoři si konkurují o malé množství herbivorů
Většina modelů tvrdí, že stabilita s rostoucí složitostí společenstva klesá – ale nemusí mýt pravdu.
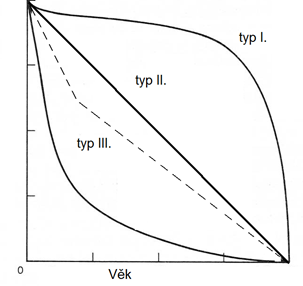
Typ I. – typický pro lidi, zvířata v ZOO, nebo listy na rostlině
Typ II. – mortalita stejná ve všech věkových třídách
Typ III. – např. ryby s miliony jiker
Na ose Y logaritmy pro snadné porovnání parametrů. Z 1000 přežije 500, ze 100 přežije 50 → v lineárním grafu se liší, v logaritmickém ne.
Největší diverzitu můžeme očekávat při „střední“ intenzitě predace, protože příliš nízká intenzita nepotlačí silné kompetitory a příliš intenzivní predace může preferovanou kořist vyhubit.
Efekt predace, parazitace a nemocí na strukturu společenstva je patrně relativně méně intenzivní tam, kde jsou fyzikální podmínky nepredikovatelné, variabilní a kruté.
Působení živočichů na strukturu společenstva se neomezuje toliko na pouhé „odebírání“ organismů. Například živočichové žijící v půdě či hrabající nory vytvářejí „disturbance“, stejně tak velcí predátoři „vyrábí“ mozaiku lokálně bohatých stanovišť.
Kohorty jsou skupiny jedinců stejného stáří (narozeny ve stejném časovém úseku). Je třeba stáří umět určit (závisí na věku, velikosti a životním stádiu).
Funkční odpověď predátora je vztah popisující závislost počtu kořisti chycené predátorem na populační hustotě kořisti.
Typ 1 – lineární růst spotřeby až do maxima
Typ 2 – rychlost spotřeby se se vzrůstající hustotou kořisti snižuje
Typ 3 – při vysokých hustotách podobná typu 2, při nižších fáze zrychlení daná růstem vyhledávací účinnosti
Typ 4 – lineární odpověď – spotřeba kořisti neomezeně stoupá
Společenstvo může být definováno na různých stupních hierarchie (spol. lesů mírného pásma, organismů pod kůrou stromu, mikroorg. v žaludku srnky, …), z praktických důvodů se na zkoumaném území označuje za společenstvo jen určitá taxonomická skupina (těžko by šlo zkoumat úplně všechno).
Vlastnosti společenstev jsou souhrné a kolektivní. Vyplývají z vlastností populací a organismů tvořících společenstvo – diverzita, biomasa, produktivita. Dále zahrnují nejen organismy, ale i jejich interakce a emergentní vlastnosti (vytvoří se, až když zkoumáme celé společenstvo, z jednotlivých prvků nejdou určit).
Zdá se, že složitější společenstva jsou celkově méně stabilní. A také se zdá, že složitá, avšak křehká společenstva obývají relativně stabilnější prostředí, a jsou náchylnější k „vykolejení“ vnějším, nepřirozeným narušením více než jednodušší, pevnější společenstva, která jsou na disturbance „více zvyklá“. Pravděpodobně bude též důležitá i paralela mezi vlastnostmi společenstva a vlastnostmi jeho populací.
Ve stabilních společenstvech budou spíše K-druhy, zatímco ve variabilních podmínkách spíše r-druhy. U K-druhů se dá očekávat, že jsou odolnější vůči narušení (jsou „silné a otužené“ z kompetice), ale jakmile už narušeny jsou, mají malou pružnost. U r-druhů naopak, nechají se snáze „vykolejit“, ale rychle se přizpůsobí.
Počet druhů je jeden z nejjednodušších parametrů. V praxi to není tak jednoduché. Záleží na počtu a velkosti vzorků.
Indexy diverzity
Rank-abundance udává rozložení početnosti druhů v rámci společenstva. Nejvyšší denzita je, když je vyrovnaný počet jedinců všech druhů → ale to trochu nejde.
Broken-stick effect – pár druhů dominuje, většina je zastoupena středně, pár je vzácných.
Podoba společenstev v prostoru
Výskyt parazita často rozhoduje o bytí a nebytí hostitele v konkrétní lokalitě – až 50 % ptactva na Hawaiských ostrovech zmizelo kvůli parazitům.
Predátoři nežijí izolovaně a jejich chování je ovlivněno přítomností jejich predátorů. Konkurence o zmenšující se zásoby potravy. K poklesu může dojít i jen díky vzájemným reakcím mezi predátory.
Druhy mají přibližně stejnou schopnost osídlit nově uvolněné plochy a poté jí bránit před jinými jedinci.
Volné plochy se tvoří neustále a nahodile → malá pravděpodobnost, že bude nějaký druh kompetitivně vyloučen – říká se tomu konkurenční loterie → jakmile nějaký organismus uhyne, jeho plocha se okamžitě otevřé nové invazi pro jedince libovolného druhu. Není tu žádná r/K strategie → nedá se očekávat sukcese.
Původní kolonizátor obvykle na území nevydrží.
Dochází k nově vzniklým uvolněným plochám, kde se střídají druhy v kontextu svých strategií. Disturbance v těchto společenstvech většinou vedou k předpověditelné sekvenci druhů – vrací stanoviště do dřívějších sukcesních stádií. Vznik „mezer“ – častý jev, hlavně pro přisedlé organismy, např. slávky a různě velké mezery uvolněné mezi nimi → invaze jiných druhů. Stejné to je i u vegetace spálené močí nebo kolem nor hlodavců.
Kompetici často ovlivňuje více počet modulů než počet genet. Zdroje jsou omezeny prostorem a konkurenty jsou nejbližší sousedi.
k hodnoty = killing values
k = log (počáteční denzita(B)) – log (finální denzita(A))
podle vzorečku teda k = log(B/A)
k se zvyšuje se zvyšující se mortalitou
jak se k mění s logB, vypovídá o charakteru vnitrodruhové kompetice
Pružnost – rychlost navrácení se do původního stavu
Odolnost – schopnost vyhnout se/odolat narušení
Lokální × globální stabilita – reakce na malé × velké narušení
Křehkost × pevnost – křehké v rámci úzkého rozmezí nějakých podmínek, robustní v širokém rozmezí podmínek
většina modelů tvrdí, že stabilita s rostoucí složitostí společenstva klesá – ale nemusí mýt pravdu
Do denzity T mají všichni dost, ale po překročení hodnoty T se rozdělí na ty, kteří mají dostatek a přežijí, a ty, co nemají nic a pojdou.
Mortalita odebírá jen nadbytečné → přesně kompenzuje.
Vnitrodruhová kompetice může na konkrétní jedince působit jinak → s intenzitou kompetice se zvyšuje variabilita jedinců v populaci (např. velikost).
Kompetice listů o světlo a kořenů o vodu – o světlo vede k větší variabilitě než o vodu, i když ta má větší vliv na biomasu.
Do denzity T mají všichni všeho dost, ale po překročení mají sice všichni pořád stejně, ale zároveň málo → všichni pojdou
Se zvyšující se denzitou se zvyšuje i fitness.
Kooperace – např. některé druhy ptactva jsou schopny společně bránit hnízda před predátorem → větší kolonie mají vyšší přírůstky (ovšem do určité hranice).
Alleeho efekt – ukazuje působení negativní kompetice v celém měřítku denzit → nízká denzita = nízká fekundita – může mít problém např. s vyhledáním partnera.
Teritorialita je jedna z nejdůležitějších ba nejběžnějších forem asymetrické kompetice. Vyskytuje se tam, kde je aktivní interference mezi jednotlivci.
Kompetice typu contest (prostředí má zdroje jen pro určitý počet jedinců). Kdo se neumístí, ten se nerozmnoží a jeho fitness je 0.
Při uvolnění místa je hned toto místo obsazeno novým jedincem. Teritorium je udržováno (hájeno) fyzickým násilím i jemněji (značkováním). Jedinci musí mít z teritoria zisk, který kompenzuje ztrátu spojenou s hájením.