Created by Kamila Klavíková
Kombinace zápisků z různých přednášek. Vytvořené jako příprava ke státnicím.
DNA a RNA (kyselina deoxyribonukleová a kyselina ribonukleová).
Jediný rozdíl mezi nimi je v ribóze.
lineární polymer, dvoušroubovice vzájemně antipararelních vláken. Tato struktura umožňuje semikonzervativní mechanismus replikace. Stabilita dvoušroubovice zajištěna vodíkovými můstky, stacking interakcemi.
Stacking interakce vznikají splýváním oblastí výskytu delokalizovaných π elektronů aromatických kruhů sousedících bazí, polaritou fosfátových skupin.
Schopnost reverzibilní separace vláken – denaturace a renaturace, možnost hybridizace (komplementární spojení 2 vláken z různých zdrojů – RNA a DNA). Jak ochotně DNA denaturuje závisí na obsahu GC (více GC → menší ochota denaturovat – 3 vodíkové vazby mezi G a C oproti 2 mezi A a T).
minor a major groove
Jsou definované vzhledem N-glykosidické vazbě. Střidají se. Jsou určené rozpětím ribóza-fosfátových skupin. Ve velkém žlábku je možno rozlišit všechny typy párů bazí (A-T, T-A, C-G, G-C).
u purinu na té straně, kde je N3 a C4; u pyrimidinu C2
celá druhá strana, ve velkém žlábku je mnohem větší množství substituentů – ty jsou rozpoznávané regulačními bílkoviny, které se vážou na DNA v oblasti velkého žlábku
V malém rozdíl mezi (A-T + T-A) a (G-C + C-G). DNA vazebné proteiny proto rozlišují sekvence DNA přednostně na velkém žlábku. Rozlišení podle přítomnosti následujících skupin vyčuhujících z dvojice bazí do žlábku: A – akceptor vodíkové vazby (vodíku), D – donor vodíkové vazby (vodíku), M – hydrofobní metylová skupina, H – nepolární vodík. Rozměry žlábku mohou vykazovat lokální změny, baze mohou být vrtulovitě zkrouceny → DNA není perfektní šroubovie.
Topoizomerázy jsou enzymy měnící topologii DNA. Odstraňují pozitivní nadobrátky vznikající před replikační vidličkou. RNA primery jsou na konci replikaci nahrazeny DNA polymerázou I a nicky jsou zaligovány DNA ligázou.
Typ I produkuje ssDNA zlom (nick). Uchovávají energii ve formě fosfotyrosinu (fosfát z nukleotidu se z jedné strany naváže na tyrosin). Reakce je reverzibilní (fosfát se po určité době spontánně naváže zpět na nukleotid). Reakce tedy nepotřebuje přísun energie ve formě ATP.
Typ II produkuje dsDNA zlom. Jsou aktivovány navázáním ATP a dimerizací v místech překřížení chromozomů. Vytvoří zlom v jednom vlákně DNA, protáhnou druhé vlákno DNA a uzavřou zlom. Hydrolýza ATP je pak vrátí do původniho stavu, ve kterém mohou vyhledat další křižující se vlákna. Topoizomerázy II jsou mimo jiné schopné oddělit propletené cccDNA (zásadní při replikaci prokaryot).
Důležité – když helikáza rozvíjí DNA, vzniká před ní nadvinutí (nadobrátka). Topoizomeráza typu I rozštípne jedno vlákno DNA, což umožní jeho otočení vůči druhému vláknu a odstranění nadvinutí.
Názvosloví bazi a umět struktury.
Nukleosid. Báze + cukr (adenosin, guanosin, thymidin, uridin, cytidin).
Nukleotid. Tvoří primární strukturu. Je to spojení nukleosidu a kys. fosforečné (ATP. cAMP, GDP...)
Báze propojené přes (deoxy)ribózofosfátovou kostru. Polarita podle 5' a 3' konce ribózy. Báze mohou tautomerizovat – adenin a cytosin na iminoformu, guanin, thymin a uracil na enolformu.
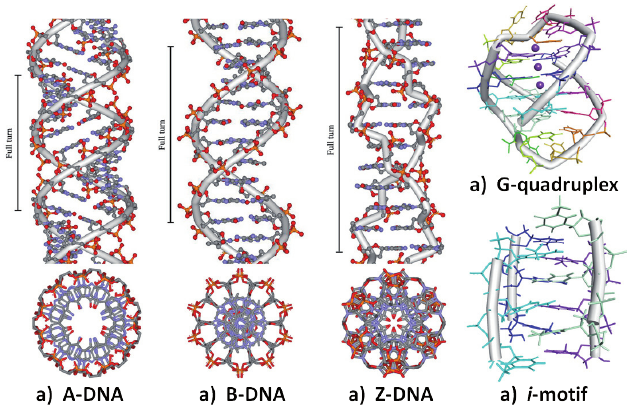
A konformace. 11 bp na závit. Pravotočivá. Krátká a silná. Velký žlábek hluboký a úzký. Malý žlábek široký a mělký.
B konformace. 10 bp na závit. Pravotočivá. Dlouhá a tenká. Velký žlábek široký (> 180°). Malý žlábek úzký (< 180°). V této konformaci se DNA převážně nachází.
Z konformace. 12 bp na závit. Levotočivá. Protáhlá cik-cak struktura. antiC, synG glykosidické vazby. Jen jeden žlábek. Druhá struktura není žlábek, ale jen vnější povrch. Vyskytuje se zejména tam, kde se střídá purin a pyrimidin. Zpravidla se jedná o GC bohaté oblasti.
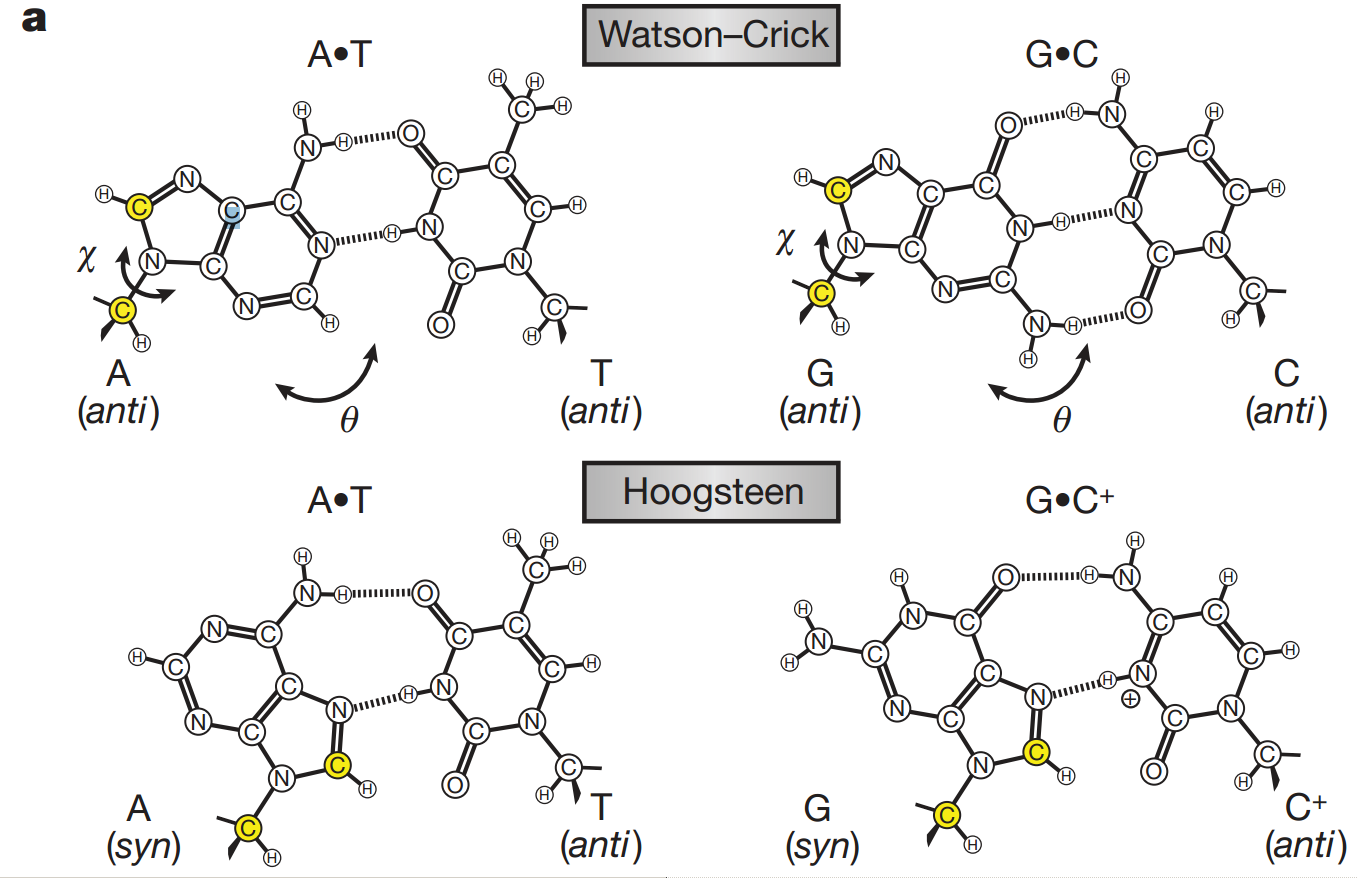
N-glykosidická vazba mezi bazí a deoxyribózou má možnost rotace, proto je možná syn a anti poloha báze vůči ribóze. N-glykosidická vazba je vazba vznikající mezi dusíkem báze a C1' uhlíkem ribózy.
Je popisovaná torzními úhly c zahrnujícími atomy O4’-C1’-N1-C2 u pyrimidinů a O4’-C1’-N9-C4 u purinů.
~ 0° (+/- 30°) = syn = cis
~ 180° (+/- 30°) = anti = trans
Pozn.: syn konformace lze uměle vyvolat např. bromací
Báze mohou tvořit vodíkové vazby s více než jen jedním partnerem → H DNA triplex, G kvadruplex (G-kvartet).
Nestandartní párování antikodónů na kodóny – U v 1. pozici tRNA antikodónu umí číst A i G v 3. pozici kodónu, G v 3. pozici antikodónu umí číst U i C v 1. pozici kodónu, inosin čte C, U, A.
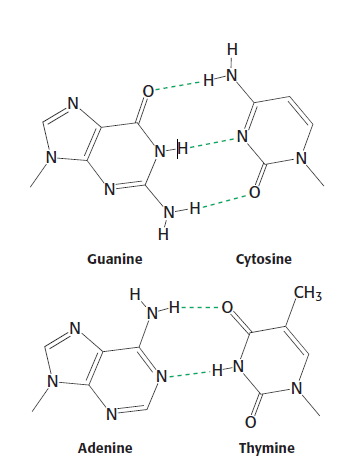
Dva řetězce nukleových kyselin spolu interagují na základě komplementárního párování pomocí vodíkových můstků.
V páru je vždy jeden purin a jeden pyrimidin. Párování poskytuje jistou symetrii. C1' uhlíky jsou v obou případech přibližně stejně daleko od sebe.
Páruje spolu adenin s thyminem (2 H-můstky) a guanin s cytosinem (3 H-můstky).
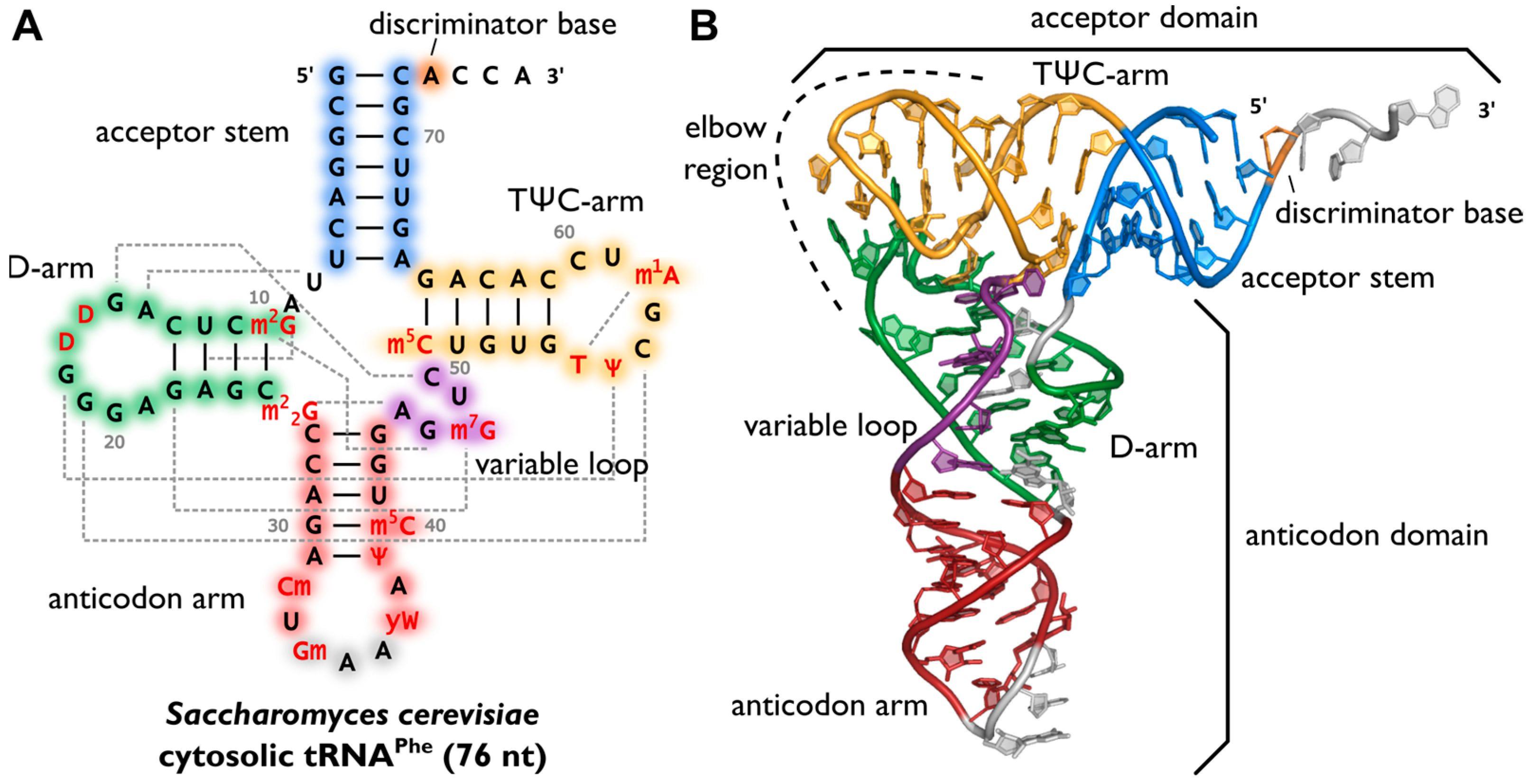
75 – 95 nukleotidů, obsahuje několik nestandartních bazí. Sekundární struktura jetelovitého listu. 4 ramena:
Akceptorové – zahrnuje 5‘ i 3‘ konec, na 3‘ konci se přes OH skupinu navázanou na C2‘ nebo C3‘ribózy posledního nukleotidu (adenylát v sekvenci CCA) váže aminokyselina
Pseudouridinové rameno (TψC) vytváří smyčku obsahující pseudouridin
Dihydrouridinové rameno (DHU) vytváří smyčku obsahující dihydrouridin
Antikodonové rameno vytváří smyčku, ve které lokalizován antikodon – triplet komplementární ke kodonu na mRNA
Vodíkové vazby ribonukleotidů tRNA, zejména na dihydrouridinovém ramenu, vedou i ke vzniku terciární struktury tRNA, důležité při proteosyntéze.
Analogy bází se začleňují do DNA během replikace.
Př. 5-bromuracil (analog T), 2-aminopurin (analog A).
Hydroxylační látky např. hydroxylamin mění cytosin na hydroxylamincytosin.
Interkalační látky se vmezeřují nebo zakliňují do DNA, čímž způsobují posunové mutace.
Př: ethidiumbromid, akridinová oranž, proflavin.
ROS (reactive oxygen species) jsou reaktivní formy kyslíku, které poškozují DNA, lipidy i proteiny. K jejich produkci dochází v mitochondriích. Asi 2-3 % kyslíku v mitochondriích jsou redukovány na ROS.
Odstraňují se pomocí superoxid dismutázy (superoxidový radikál → H2O2), katalázy (H2O2 → H2O), glutathion peroxidázy.
Neexistuje vždy korelace mezi rychlostí metabolismu a délkou života (rychlejší metabolismus…– kratší život). Existuje celá řada výjimek jako ptáci, vačnatci, netopýří… Ač je tělo rypoše vystaveno velké míře oxidativního stresu, dožívá se v zajetí 28 let. Tedy musí mít jiné mechanismy.
Př.: peroxidy, superoxidy, hydroxylový radikál, singletový kyslík.
Alkylační látky přidají alkylovou skupinu, čímž způsobují substituce (tranverze a tranzice) a chromozomové aberace.
Př. EMS, MMS ⇒ alkylace G v poloze 6 ⇒ párování s T ⇒ změna GC na AT
Kyselina dusitá nahrazuje amino skupinu keto skupinou, tomuto procesu se říká oxidativní deaminace. Působí na A, C, G.
loss of heterozygosity (LOH)
Mutace alely určitého genu v somatické buňce, která již nese na homologickém chromozómu mutovanou alelu téhož genu.
Změna exprese, NE produktu.
down mutace → snížení transkripce + zvětšení vzdálenosti od START nukleotidu
up mutace → zvýšení transkripce; zmenšení vzdálenosti od START nukleotidu.
Netýkají se DNA; většinou při transkripci; projev obvykle slábne s časem.
Mutace v 3´sestřihovém místě → sestřihový aparát toto místo přeskočí a dojde k odstranění exonu z transkriptu
Mutace v 5´sestřihovém místě → zachován intron v transkriptu
Ve stejném místě jako byla mutace přímá.
Pravá reverze. Obnova mutované skevence nukleotidů na původní. Opačná reverze – změna kodonu na kodon synonymní k původnímu kodónu (před pímou mutací).
Částečná reverze. Změna kodónu na nesynonymní kodón, leč takový, že dojde k úplné či častečné obnově funkce s novou AK oproti mutaci přímé.
Vliv na expresi jednoho sousedního genu; geny lokalizovány na jedné straně mutantního genu.
Mutace v genu, který kóduje produkt účastnící se regulace transkripce jiného genu/mutace v regulační oblasti genu.
Submikroskopičtí obligatní celulární parazité. Nemají vlastní energetický metabolismus, nerostou, vznikají spojením jednotlivých komponent. Dělit se dokáží jen v buňce.
Virová částice se nazývá virion. Minimální výbava virionu – dědičná informace (RNA / DNA) a virová kapsida (proteinový obal NK). Dále může virus obsahovat další obal kolem kapsidy a různé enzymy a proteiny.
Virové genomy jsou malé a komplexní. Genom je často složen pomocí virových histonů a asociován s proteiny kapsidy. Genom musí být čitelný pro replikační a transkripčně translační aparát buňky.
Kapsida je fyzikálně a chemicky odolná. NK kóduje i další potřebné proteiny.
Všechny možné variace způsobů replikace (dsDNA, ssDNA, dsRNA, (+)(-)ssRNA).
Supresorová mutace. Maskuje účinek přímé mutace; intragenová (mutace v témže genu, ale na jiném místě) X intergenová (mutace v jiném genu než přímá mutace)
Enhancerová mutace. Opak supresorové; zvýšení intenzity mutačního fenotypu; často letalita.
Buněčný typ: somatické buňky / buňky zárodečné linie
Typ chromozomu: autozomální / gonozomální
Molekulární změna: substituce / inzerce / delece
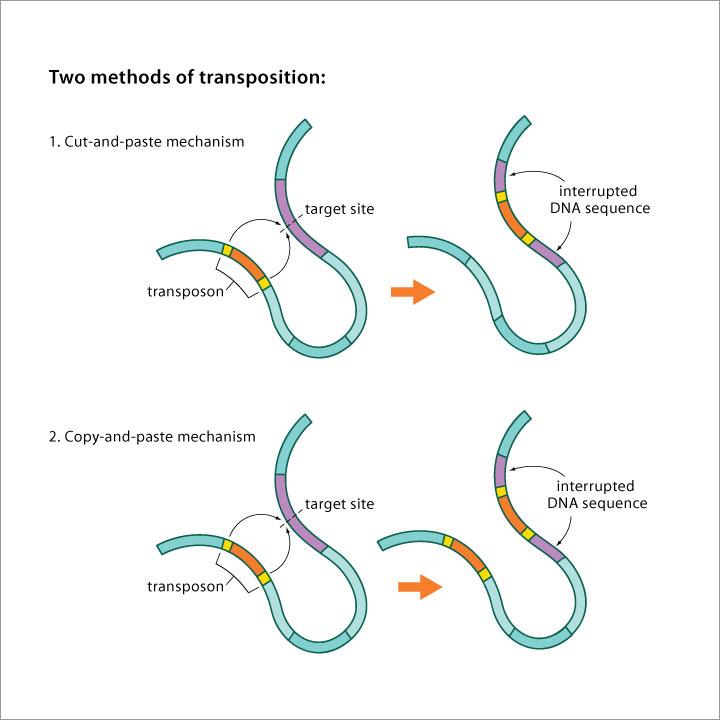
Nesou informaci nutnou pro svůj vlastní pohyb (geny kódující enzymy umožňující transpozici).
Neschopnost pohybu bez účasti autonomních elementů. Mohou je však přenést. Obvykle to bývali autonomní elementy, které ztratily gen/y (či mají poškozený gen/y) pro enzym umožňující transpozici.
mobilní genetické elementy, transpozony, rozptýlené repetice
Transponovatelné elementy jsou úseky DNA schopné změnit svou pozici / pohybovat se v rámci genomu jednoho organismu (v širší def. i mezi organismy). Jsou to rozptýlené repetitivní sekvence.
Vyskytují se u archeí, bakterií, eukaryot i virů (Pandoraviry).
Transpozony se často šíří i horizontálně. V evoluci se střídají periody, kdy jsou TE aktivní a množí se, s periodami klidu.
Zpravidla se v genomu vyskytují pouze malá množství celých nepoškozených transpozonů. Většinu tvoří nefunkční, zkrácené či jinak poškozené sekvence transpozonů. Ale i poškozené se mohou kopírovat, pokud v genomu je alespoň několik nepoškozených kopií, které dodávají potřebné enzymy. Transpozony mohou tvořit významnou část genomu (1000 až 10.000 kopií, člověk 50 %, kosatec 98 %) .
Objev transponovatelných elementů na Ac/Ds elementech kukuřičných zrn (aktivátor - autonomní, disociátor - neautonomní).
Dominantní. Pokud projevena pozdě v ontogenezi, přenos do dalších generace.
Recesivní. V populaci zachovány hlavně u heterozygotů. Neobvyklé štěpné poměry v potomstvu F2 generace.
Auxotrofní. Neschopnost syntetizovat určitou důležitou živinu.
Teplotně senzitivní. Produkt labilní za určitých teplot.
Životaschopnost pouze za přítomnosti supresoru.
Ztrátová mutace. Snížení funkce (= leaky mutace) + ztráta funkce (= null mutace) mutovaného genu.
Zisková mutace. Zvýšení funkce; větší množství produktu; zisk nové vlastnosti
Pro transpozici využívají reverzní transkriptázu. Copy-and-paste přes RNA intermediát. Představují značnou část genomu (u člověka 42 % / zatímco DNA transpozóny jen 2-4 %).
LTR retrotranspozóny. Na koncích mají LTR (long terminal repeats) – dlouhé přímé repetice. Velmi časté u rostlin, vzácnější u živočichů. Podobné retrovirům. Pravděpodobně vznikly z retrovirů. Jsou to pozůstatky dávných virových infekcí.
PLE retrotranspozony. PLE (penelope-like elements) na koncích mají LTR (můžou být invertované), tvoří TSD(target site duplication) – krátkou přímou repetici. Kódují endonukleázu.
Končí AT bohatou oblastí.
LINES (long interspersed nuclear elements). Délka až několik kbp. Velmi časté u živočichů, vzácnější u rostlin.
SINES (short interspersed nuclear elements). Nevirové neautonomní retrotranspozóny. Nekódují reverzní transkriptázu. Pro transpozici využívají reverzní transkriptázu jiných elementů (většinou LINES). Tvoří TSD. Na jednom konci sekvence bohatá na AT o délce 100-500 bp.
Somatické mutace. Projev u potomků somatické buňky; nepřenáší se gametami do potomstva; ztráta heterozygozity = mutace alely genu v somatické buňce, která už nese na homologickém CH mutovatou alelu téhož genu (retinoblastoma u člověka); dominantní mutace se projeví ve fenotypu
Gametické mutace. Přenos gametami do potomstva; dominantí mutace projev ve fenotypu potomstva
Indukované mutace sou způsobené mutageny, což jsou látky, faktory vnějšího prostředí, které indukují vznik mutací. Jsou různé povahy, biologické, fyzikální a chemické.
Bodové mutace. Záměna dvou nukleotidů (inzerce, delece, transverze – různý typ báze, tranzice – stejný typ báze)
Řetězcové mutace. Záměna celého úseku DNA (duplikace, inzerce = zmnožení řetězce DNA, delece = vystřihnutí úseku DNA, translokace = vzájemné vyměnéní úseku DNA, inverze = určitý úsek DNA je z chromozomu vystřihnut a vložen do stejného místa v opačné orientaci – může způsobit reprodukční bariéru a tím speciaci)
Chromozomální mutace. Translokace velkého rozsahu vznikající při mitóze. Následky drastické – mládě se ani nenarodí, nebo nemoci typu Downův syndrom.
Genomové mutace. Probíhají na úrovni chromozomů či chromozomových sad v důsledkem poruch v průběhu buněčného dělení (aneuploidie, polyploidie, autopolyploidizace, aloploidizace)
Reorganizace DNA v důsledku rekombinací mezi 2 kopiemi transponovatelného elementu. Vznikají delece, inverze. Závisí na tom, zda jsou kopie přímé nebo invertované. Dochází k zvětšování genomu.
Záleží na místě inzerce. Možný vliv i po excizi elementu v rámci cut-and-paste transpozice. Transponovatelné elementy mají evolučně získanou tendenci se inzertovat do safe heavens, nekódujících částí genomů, kde jejich inzerce nepoškodí hostitele.
Část neutrálních mutací je slabě negativní. V obrovské populaci by se chovaly negativně.
Z plazmidu do plazmidu, či z plazmidu do bakteriálního chromozomu.
Pro transpozici nevyužívají reverzní transkriptázu. Jsou lemované obrácenými repeticemi.
Transpozice cut-and-paste. Při transpozici vznikají dvouřetězcové zlomy.
TIR elementy. Mají na koncích invertované repetice (TIR). Transpozice transpozázou.
Ac/Ds elementy u kukuřice.
P elementy. Nejvýznamnější elementy u Drosophily. Jejich kódování podléhá alternativnímu sestřihu – buď transpozáza nebo represorový protein.
Transpozice copy-and-paste, ale ne přes RNA intermediát. Při transpozici vznikají jednořetězcové zlomy (vznik kointegrátu).
Helitron elementy. Na koncích nemají TIR a netvoří TSD. Zejména u rostlin.
Maverick elementy. Na koncích TIR. Jsou velké (až 20 kbp). Nezjištěny u rostlin. Specifický mechanismus transpozice.
změny smyslu (vlivu na translaci)
Synonymní mutace. Substituce v jedné bázi, která nemění kódovanou AK.
Missense mutace. S chybným smyslem. Změna kodonu pro jednu AK za kodon pro jinou AK.
Nonsense mutace. Nesmyslná substituce vedoucí ke vzniku stop kodonu na místě kodonu pro AK.
Spontánní mutace vznikají přirozeně bez známé příčiny.
Při deaminaci dochází k nahrazení aminoskupiny (-NH2) ketoskupinou.
Cytosin. Hydrolýza na uracil.
5-methyl-cytosin. Hydralýza na thymin. Místo nejčastějších mutací u obratlovců.
Adenin. Vzniká hypoxantin, který se raději váže na cytosin. Vzniká tedy v podstatě GC pár.
Guanin. Vzniká xantin, který se páruje s thyminem. Nový pár je v podstatě AT.
Ztráta purinové / pyrimidinové báze a vznik apurinického / apyrimidinového místa (AP místa).
Př: ztráta purinové báze (A, G) ⇐ přerušení glykozidické vazby mezi deoxyribózou a purinem
Sklouznutí (slippage) je sklouznutí DNA polymerázy během replikace, lokální denaturace a následná reparace. Dopředné sklouznutí – delece, zpětné sklouznutí – adice. Může k němu dojít kdekoliv v DNA. Nejčastěji se tak děje v repetetivních sekvencích.
Báze je pozměněná změnou pozice vodíkového atomu. Pozmění se způsob tvorby vodíkových můstků, čímž se změní párování. Stabilní formy bazí – keto (G, T) a amino (A, C), méně stabilní enol a imino.
rentgenové záření, γ záření a kosmické záření…
Ionizující záření navozuje strukturní změny chromozomů v důsledku dvojchromatidových zlomů. Proniká v různých vzdálenostech do tkání. Využívá se v klinické praxi.
UV záření způsobuje vznik thyminovch dimerů (mohou vznikat i TC, CC dimery) a cytosinhydrátu. Proniká do tkání pouze povrchovou vrstvou buněk. Je snadno absorbováno organickými molekulami. Nezpůsobuje ionizaci. Vede k exitaci elektronů do vyšší energetické hladiny, čímž způsobuje vyšší reaktivitu molekul.
Počet mutací v dané pozici za jednotku času všech členů populace.
Výskyt dané mutace vyjádřen jako podíl mutantních jedinců v populaci. Vyšší frekvence mutací u samců (vyšší selekční tlak na X chromozom a vyšší počet buněčných dělení během spermatogeneze).
Klasický darwinismus je založen na postupné kumulaci mikromutací, tedy mutací, které způsobují malé změny oproti původnímu stavu.
Existují ale i makromutace, jejichž vlivem se stavba či fungování těla změní výrazným způsobem (třetí ruka, jedna noha...). Je však otázka, zda by takovýmto způsobem mohly vzniknout některé zásadní nové orgány jako např. křídla apod. Makromutace existují, ale jejich role v evoluci je nejistá. Jedině díky makromutacím můžou vzniknout mezidruhové rozdíly.
Opraví téměř každé poškození, např. dané navázáním karcnogenu na bázi DNA, nebo pyrimidinový dimerem.
Skenuje se DNA a hledá se distorze šroubovice, nikoli změna primární struktury (na jednotlivých bazích).
U E. coli 4 proteiny Uvr (UV repair?) zodpovědné za tento systém. UvrA*UvrB skenují DNA, za detekci je zodpovědná UvrA, po detekci UvrB rozvine DNA v místě distorze. UvrC (endonukleáza) pak štěpí 8 nukleotidů od leze na 5' straně, 4-5 nukleotidů od leze na 3' straně. UvrD (helikáza) odstraní poškozený úsek. Mezera je zaplněna DNA polymerázou a zalepena DNA ligázou.
U eukaryot až 25 proteinů – XP geny, jejich mutace vede ke zvýšené citlivosti vůči UV → choroba Xeroderma pigmentosum. Transcription-coupled DNA repair – zabrzdí li se RNA polymeráza na chybě v DNA, přijdou opravné proteiny a poškození opraví → RNA polymeráza může fungovat jako senzor poškození DNA.
Telomeráza u eukaryot. Při ukončení replikace není možné doplnit poslední nukleázou odstraněné RNA primery; telomerasa nepotřebuje templát a funguje mechanismem reverzní transkriptázy; nasedá na 3' přesah a prodlouží konec; řeší problémy konců telomer a brání jejich zkracování při replikaci.
Myšlenka možnosti přechodu jednoho druhu v jiný prostřednictvím přepínání alternativních ontogenetických programů. Př. když se pšenici nedaří, začne se měnit na žito, kterému by se za daných podmínek dařilo lépe.
Tato myšlenka není úplně zavrhnutelná, ale v rámci celého lysenkismu (falšování, ideologie) je mimo.
Fotolyáza opraví pyrimidinové dimery. Odstranění metylace methyltransferázou.
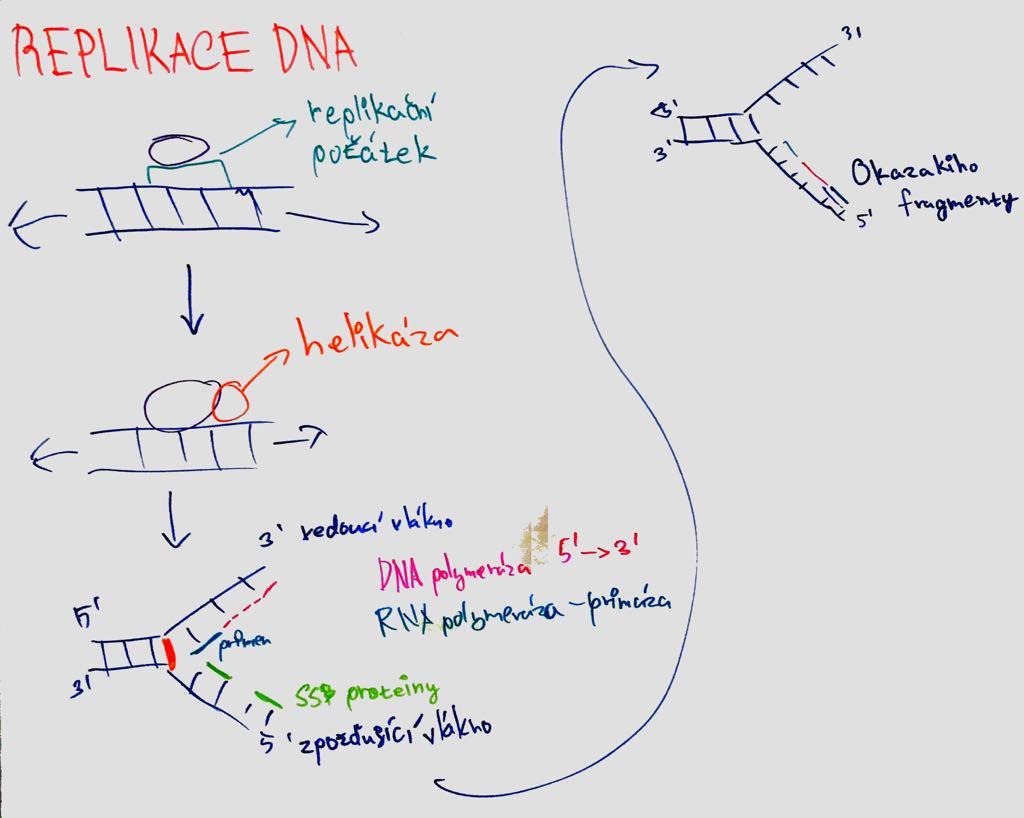
semikonzervativní = nová molekula z 1 starého a 1 nového vlákna; obě řetězce jsou komplementární a mohou sloužit jako templát.
replikační aparát = soubor proteinů pro replikaci;
replikační počátky = ori = místa, kde se dvoušrobovice poruší první (tato místa jsou rozpoznána iniciačními proteiny; bohaté na AT; u bakterií je genom kruhový a tedy 1 počátek);
replikační vidlička = začátky replikace ve tvaru Y, kam se vážou proteiny repl. aparátu (pohyb ve směru replikace a rozvíjejí šroubovici za syntézy nového vlákna; v 1 ori se tvoří dvě a pohybují se od sebe);
DNA polymeráza = enzym katalyzující polymeraci řetězce DNA – vkládá deoxyribonukleotidy a prodlužuje řetězec (tvar ruky; palm doména kontroluje párování; finger doména uzavře dNTP v katalytickém místě; thumb doména drží syntetizovanou DNA; katalyzuje připojení nukleotidů na 3´konec; v aktivním místě jsou Mg ionty; NTP dodává energii; má opravnou funkci = proofreading – před přidáním nukleotidu kontroluje předchozí pár); syntéza ve směru 5´- 3´; vedoucí řetězec = leading strand (syntéza kontinuálně) X opožďující = lagging strand (diskontinuálně;
Okazakiho fragmenty = primery – nukleáza odstraní RNA primery, DNA Pol je nahradí DNA a ligáza spojí));
primery = krátké úseky RNA (DNA POL neumí začít syntézu sama; tvorba primázou);
helikáza = rozvíjí ATP za spotřeby ATP; SSB proteiny = ochraňují 1vláknovou DNA před spojením;
sliding clamp proteiny = svorkové (vážou DNA POL na templát); replisom (komplex proteinů podílejícíh se na replikaci v místě vidličky)
Mutace jsou změny ve struktuře genetického materiálu, při nichž se mění smysl genetické informace, aniž by byla porušena syntaktická pravidla jejího zápisu. Jako mutace označujeme změny sekvence DNA.
Je třeba rozlišovat mutace, respektující pravidla zápisu do DNA, a poškození DNA. Mutace jsou největším zdrojem genetické variability.
Genové mutace. Postihují jen jeden gen.
Chromozomové mutace. Změna struktury chromozomů, pořadí genů, polohy centromer.
Genomové mutace. Celková změna genomu (polyploidizace, aneuploidizace).
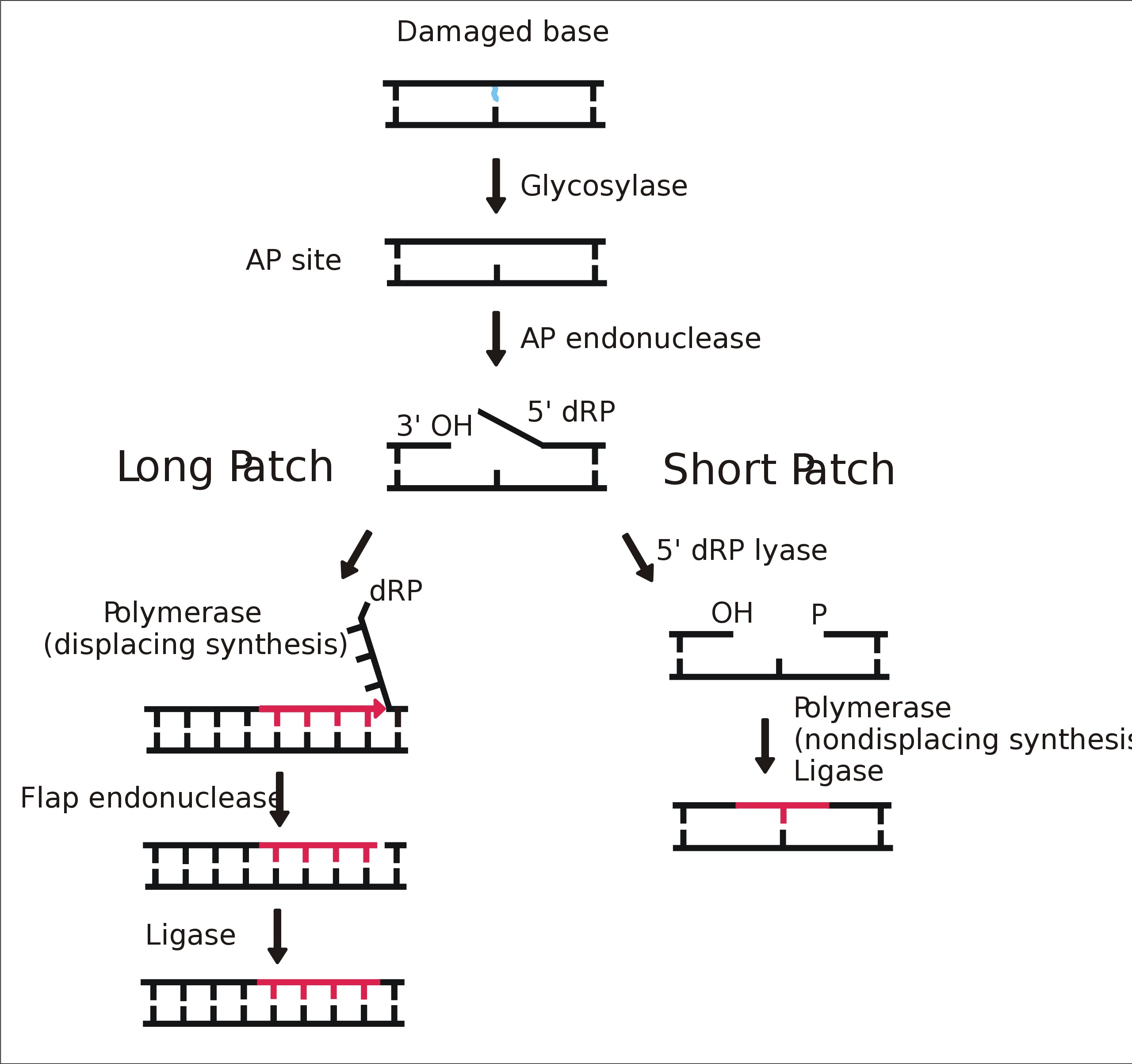
Oprava podle nepoškozeného vlákna (závislost excizní reparace na ds charakteru nukleové kyseliny). 3 obecné kroky:
Fotoreaktivace. Enzym DNA fotolyáza po osvětlení DNA UV zářením opraví pyrimidinové dimery. Využívá energii viditelného světla.
Odstranění metylace. Metyltransferáza přenese metylovou skupinu z 6-O-metylguaninu na cystein, energeticky velmi náročné.
Místa výskytu mutací nejsou v řetězci DNA rozmístěna rovnoměrně – častěji k nim dochází na horkých místech (hot spots). Změny frekvence mutací pozorujeme i v čase, kdy jich v některých místech evoluce vzniká podstatně více – není úplně objasněno proč – sekvenční motiv (AT sekvenční motivy), vnější podmínky, pohlavní rozdíly (samci mutují častěji)… Jsou také místa, kde sice nevzniká více mutací, ale kde se mutace snáze fixují.
Některé organismy dokážou ve stresových podmínkách začít mutovat rychleji, což jim zvýší šanci na přežití. Někteří autoři se domnívají, že nejen mutují rychleji, ale i cíleně, např. trypanosoma dokáže vytvářet mnoho různých variant povrchových proteinů a dělá to záměrně v určitý čas a za určitých podmínek.
Liší se mezi pohlavími. Častěji mutují samci – při vzniku spermií mohou zkoušet evoluční novinky. Samice jsou konzervativnější, má jich méně a nevyplatilo by se jí riskovat.
Role mutátorů – patogenní bakterie mají v populaci hodně mutátorů, snaží se tak najít optimální virulenci.
kruhová molekula DNA; 1 ORI;
a) replikační vidlice oběma směry a nasedá na DnaA protein – DnaB helikáza rozvolňuje šroubovici – primáza nasyntetizuje primer – DNA POL III prodlužuje řetězec; primozom = komplex helikázy a primázy se pohybuje kolem DNA; DNA POL III prodlužuje primery – DNA POL I vymění RNA za DNA – ligáza je spojí; terminace replikace v terA a terB;
b) valivá kružnice; jednovláknové přerušení kružnicové molekuly – replikuje se druhé neporušené vlákno – vznik produktu, kde jsou za sebou umístěny kopie plazmidu – rozstříháno na správnou velikost a zaligováno
Samostatná existence cílených mutací by ale nestačila k lamarckistické evoluci. Naprostá většina mutací je náhodná.
První překážkou lamarckistické evoluce je neexistence zpětného toku z proteinů do DNA.
Druhou překážkou je Weismanovská bariéra mezi germinální a somatickou linií.
Třetí překážkou je fakt, že genetická informace není přesný popis struktury.
Jedním ze směrů popírajících darwinismus je mutacionismus.
Jeho zastánci tvrdili, že nejdůležitějším zdrojem mutací jsou reparační procesy.
Nebyli tak ale schopni objasnit vývoj adaptivních znaků. To vysvětluje mechanismus přirozeného výběru, který je nepostradatelný pro šíření výhodných mutací.
Mismatch repair systém opravuje špatně zařazené nukleotidy při replikaci. Opravuje chyby, které neopravila DNA polymeráza.
Systém musí rozpoznat, kterou bázi při nesprávném párování vyměnit, musí vědět, které vlákno sloužilo jako templát při proběhnuvší replikaci.
U gramnegativních bakterií je templátové vlákno metylováno, nové vlákno ještě ne. U ostatních prokaryot a eukaryot není mechanizmus jasný, u eukaryot je ne-templátové vlákno pravděpodobně odlišeno přítomností nicků. Ty se objevují i ve vedoucím vlákně. Zda jsou tam uměle zavedeny, není jasné. Proteiny u E. coli– MutS, MutL, MutH, svorkový protein PCNA
Část chybně spárovaných bazí opraví DNA polymeráza – palm doména kontroluje párování právě připojeného nukleotidu, pokud zjistí chybu, tak se primer*templát komplex uvolní z polymeračního místa (thumb a finger) a jde na opravné místo na palm doméně, kde nastupuje 3‘→5‘ exonukleázová aktivita palm domény.
Oprava dvouvláknových zlomů (vznikají při ionizačním záření, vlivem oxidačních látek), kdy mezi vlákny zlomu dojde ke ztrátě nukleotidů.
error prone repair
trans-lesion DNA synthesis: Když DNA polymeráza narazí neopravenou chybu, disouje i se svorkovým proteinem. Na její místo nastupuje translesion DNA polymeráza, která překlene místo poškození, ale generuje chyby (není tak závislá na párování bazí).